РЕФЛЕНСЫ
РЕФЛЕНСЫ (от лат. reflexio—отражение}, автоматические двигательные реакции в ответ на внешнее раздражение. Термин Р. заимствован из области физ. явлений и имеет в виду аналогию между нервной системой, отражающей раздражение в форме двигательной реакции, и гладкой поверхностью, отражающей падающий на нее луч света (Astruc, 1743). По своей
?биол. •сущности Р. представляет собой одно из проявлений основной функции нервной системы— раздражительности. Как известно, раздражительность представляет собой свойство, присущее всякой живой протоплазме, и довольно сложные двигательные реакции в ответ на внешние раздражения наблюдаются у растений, у простейших животных и в блуждающих клетках вьющих организмов (лейкоциты, семенные клетки и т. п.). Подобные реакции, осуществляемые без участия нервной системы и обозначаемые терминами «тропизм» (изменение в положении частей неподвижного организма) и «таксис» •(перемещение всего организма), представляют собой наиболее примитивный стадий в эволюции раздражительности. Стадий этот может быть ■обозначен как период «простой раздражительности». Сущность раздражительности состоит в изменении под влиянием внешнего воздействия физ.-хим. состояния и освобождении энергии; динамическое изменение, происшедшее в месте воздействия раздражения, может вызвать аналогичное изменение в соседней области и т.о. процесс раздражения может распространяться, иррадиировать, проводиться из одного отдела организма на другой. Таким образом уже в том -стадии, к-рый мы обозначаем термином «простая раздражительность», имеется элементарная форма проводимости. Существенная особенность этого распространения раздражения в нервной ткани по сравнению с нервной системой заключается во-первых в медленности процесса распространения, а во-вторых в быстром уменьшении интенсивности раздражения по мере его удаления от места его возникновения, т. е. от места, непосредственно подвергшегося внешнему раздражению; постепенно ослабляясь, раздражение на известном расстоянии от места его возникновения сходит на-нет. В дальнейшем ходе эволюции, в процессе морфол. и фнкц. специализации и приспособляемости тканей возникает нервная система как продукт превращения эпителиальной ткани. Биол. сущность появления нервной клетки может быть определена как превращение эпителиальной клетки в смысле ее исключительного приспособления к специфической функции раздражительности и проводимости. Морфол. выражением этих специфических свойств нерв- ной клетки является наличие б. или м. длинных отростков, при помощи к-рых эти клетки приводят в сообщение различные, иногда весьма отдаленные части организма; фнкц. выражением специфических свойств нервной клетки является понижение порога, раздражения по отношению к различного рода воздействиям и способность проведения раз возникшего процесса раздражения рез замедления его распространения и без ослабления его интенсивности. Т. к. такой именно способ распространения процесса возбуждения обозначается термином «проводимость», то этот период эволюции реакций организма на внешние раздражения может быть обозначен как стадий раздражительности и проводимости. Хотя в этом стадии двигательные реакции организма на внешние раздражения осуществляются при посредстве нервной системы и хотя при этой форме реакций имеются основные фнкц. составные части рефлекса—раздражение, проведение и реакция, тем не менее здесь еще не имеется рефлекторного акта в истинном смысле этого слова. Для уяснения отличия этой примитивной формы нервной функции от настоящего рефлекторного акта и для уяснения сущности последнего необходимо иметь в виду следующее. На первом стадии своего появления в процессе филогенетической эволюции нервная система представлена, как это имеет место напр. у кишечнополостных, в виде сети, диффузно распространенной по всей массе тела; при этом, где бы на воспринимающей раздражения поверхности тела ни возникло раздражение, оно распространяется по нервной сети во всех направлениях. Если из колокола медузы вырезать спиральную или зигзагообразную полоску, то при раздражении любого конца этой полоски получается сокращение этой полоски. Из этого следует, что в примитивной нервной системе, имеющей сетеобразное строение, нервный импульс, т. е. процесс распространения возбуждения, может осуществляться во всех направлениях; другими словами, каждый из нервных стволов, составляющих эту сеть, проводит возбуждение в обоих направлениях своей оси. Такие свойства нервной системы—ее сетевидное строение и распространение импульса по всей сети при раздражении любого участка—вполне приспособлены к тем формам реакций, т. е. тем способам движений, к-рые свойственны соответствующим животным,-—именно для диффузных мышечных сокращений всей массы тела. Эта форма строения и функционирования нервной ткани обозначается термином «эквипотент-ная система». В противоположность этому у высших беспозвоночных и всех позвоночных животных не-вроны сочетаны в известные комплексы (цепи), представляющие определенную закономерность в отношении направления импульсов. Эта специфичность распространения импульсов, обозначаемая термином «динамическая поляризация», состоит в том, что импульс распространяется от дендрита клетки через ее тело к осево-цилиндрическому отростку и далее, по этому последнему—к дендриту следующей клетки и т. д.; другими словами, направление импульса центростремительно (по отношению к телу клетки) в дендрите и центробежно в аксоне. Помимо динамической поляризации нервная система высших беспозвоночных и всех позвоночных представляет целый ряд других особенностей, отличающих ее от диффузной эквипотенциальной ' системы кишечнополостных и низших червей. Особенности эти заключаются в следующем. Эквипотентная нервная система представляет «обой действительную сеть нервных волокон с несомненным переходом фибрилярных волокон из одного нерва в другой. В нервной системе высших животных такой анат. непрерывности между невронами не существует; различные невро-ны находятся здесь между собой лишь в анат.-физиол. контакте; такие области контакта между невронами, области, где концевые разветвления аксона одного неврона приходят в сообщение с разветвлениями дендрита другого неврона, обозначаются термином синапс. Синаптические связи не являются простым анат. соприкосновением между отростками клеток; синапсам принадлежит весьма существенная физиол. роль. Прежде всего следует отметить, что прохождение импульса через синапс связано с нек-рым замедлением в его распространении, что имеет весьма важное значение для высших форм двигательных актов, характеризуемых чертой выбора («волевые движения»). Далее, можно считать установленным, что синапс является фактором поляризации сочетающихся в цепь невронов, т. е. фактором распространения , нервного импульса в определенном направлении, именно в направлении от аксона одного неврона к дендриту другого. Синапсам принадлежит также повидимому существенная роль в торможении и стимулировании импульсов и в определении т. н. «конечного пути» или «последнего общего этапа» (final common path, см. ниже), т. е. направления импульса на определенный комплекс периферических невронов при наличии совместного воздействия на последние различных иннервации. В общем синапсу приписываются те особенности функции проводимости. которые свойственны нервным процессам, осуществляемым цепью невронов по сравнению с проводимостью в отдельном нервном волокне. В механизме рефлекторного акта,.в осуществлении к-рого всегда принимают участие не менее двух невронов, синапсу принадлежит т. о. существенная роль. Морфологически сущность синапса видели в наличии между невронами особой мембраны; существование последней нельзя считать вполне доказанным. Другие видят морфол. сущность синапса в том промежутке, к-рый образуется между невронами вследствие отсутствия между ними непрерывной связи. Шеррингтон (Sherrington) обозначает это пространство термином «поверхность разделения» (surface of separation); такая поверхность может обладать различными физ. свойствами, к-рыми могут быть объяснены функции синапса: задер- , живать диффузию, влиять на осмотическое дав- ' ление, движение ионов и т. п.; определяемая синапсом проводимость лишь в одном направлении могла бы быть объяснена тем, что мембрана (или поверхность разделения) обладает односторонней проницаемостью. Какова бы ни была морфол. природа синапса, можно считать общепринятым, что синапс не "есть инертная полость между невронами; за ним должна быть признана роль активного элемента нервной системы, «живой субстанции, отличающейся от остальной протоплазмы невронов» (Herrick). Все приведенные особенности функции нервной системы являются необходимыми элементами рефлекторного акта: участие в нем не менее двух (а обычно более) невронов; закономерная проводимость в определенном направлении (от рецептора к эффектору); необходимость ограничения двигательной реакции определенной группой мышц,- а не диффузная реакция всей мускулатурой тела. Все эти свойства двигательной реакции находятся в неразрывной связи с теми морфол. и фнкц. особенностями, к-рые присущи всем животным, стоящим выше кишечнополостных и простейших червей, т. е. значительной части беспозвоночных и всем позвоночным. Основным типом строения всех этих животных являются два признака: билятеральная симметрия и сегментарность, или метамерия. Строение организма по сегментарному типу предполагает известную степень самостоятельности каждого из сегментов и соответственно этому самостоятельность (изолированность) двигательных реакций отдельных сегментов или их групп, а в нек-рых случаях и одной половины тела. Рефлекторный акт в строгом смысле слова появляется на пути филогенетического развития одновременно с появлением сегментарного билятерально-симметричного строения тела; вместе с тем в своей наиболее элементарной форме Р. представляет собой сегментарную реакцию на внешнее раздражение. Т. к. вся эволюция нервной системы состоит в надстройке новых элементов (надсегментарный аппарат) над старыми, то с фнкц. точки зрения Р. является основой, над которой в процессе эволюции надстраивались более сложные механизмы нервных функций. Эти последние, т. н. над сегментарные механизмы развивались однако не за счет исчезновения простых сегментарных реакций, а за счет подчинения сегментарного рефлекторного механизма вновь вырабатывающимся аппаратам и за счет приспособления сегментарных рефлекторных актов к филогенетически более новым функциям. Поэтому механизмы примитивного сегментарно-рефлекторного аппарата сохранились и у высших животных. С другой стороны, следует признать, что сегментарный Р. представляет собой основной тип реакции на внешние раздражения и что все более высшие формы этих реакций являются не чем иным, как постепенным усложнением и превращением простого рефлекторного акта. И с этой точки зрения «все акты сознательной и бессознательной жизни по способу своего происхождения суть рефлексы» (И. М. Сеченов). Однако практически перенесение термина «рефлекс» на все проявления нервной деятельности едва ли представляет какие-либо выгоды, потому *что оно не ведет к уточнению понятия о различных формах деятельности нервной системы. Хотя весь ход эволюции двигательных функций,.начиная с тропизма безнервных организмов и кончая высшими проявлениями функций мозговой коры человека, представляет собой усложнение, превращение и совершенствование реакций живой материи, тем не менее для точного анализа различных видов этих по своей основной сущности аналогичных явлений их выгоднее не сочетать в одну общую группу, а различать их отдельные формы. С этой точки зрения под рефлексами и следует разуметь сегментарные двигательные реакции, т.е. реакции, осуществляемые нервными механизмами спинного мозга и стволовой части головного мозга, хотя эти механизмы находятся под влиянием более высоких (надсегментарных) отделов головного мозга. Ограничивая т. о. понятие о Р. двигательными реакциями сегментарного аппарата, мы можем охарактеризовать их следующими специфическими чертами, отличающими их от двигательных функций, осуществляемых корой голов но- 76№> го мозга в форме т.н. произвольных или волевых движений. Р. представляют собой двигательные акты, осуществляемые роковым образом в ответ на внешнее раздражение; рефлексы бессознательны, автоматичны; волевые движения сознательны, произвольны; рефлексы имеют врожденный характер и осуществляются по предопределенным анат. путям; волевые движения приобретаются в течение индивидуальной жизни и соответствующие им анат.-физиол. связи не являются предопределенными, но вырабатываются на пути индивидуального опыта данного субъекта; соответственно этому Р. имеют видовой характер, т. е. присущи всему виду Homo sapiens, в то время как волевые движения имеют индивидуальный характер; анатомически Р. суть реакции спинного мозга, тогда как волевые реакции осуществляются корой головного мозга; с точки зрения физиологии условных рефлексов сегментарные Р., т. е. Р. в узком смысле этого слова, представляют собой безусловные Р., в то время как волевые движения соответствуют понятию об
условных рефлексах (см.). Между Р. и волевыми движениями существуют переходные формы, что представляется вполне понятным в виду постепенности перехода от Р. к волевым движениям; поэтому существуют двигательные реакции, к-рые, относясь в главных своих чертах к одной из названных групп, имеют отдельные черты, характерные для другой группы. Так напр. нек-рые из реакций, относимых к Р., являются не врожденными, а приобретаются в течение индивидуальной жизни (напр. кожные, брюшные Р.); с другой стороны, нек-рые из заведомо корковых движений, хотя и приобретаются в течение индивидуальной жизни, но имеют видовой характер (напр. акт хождения); нек-рые из приобретаемых в течение индивидуальной жизни корковых движений вследствие их частого повторения автоматизируются в такой степени, что приближаются в этом отношении к Р. Кроме того следует иметь в виду, что нек-рые из врожденных автоматических двигательных реакций подвергаются на пути онтогенетического развития, в процессе приспособления к новым функциям, различным превращениям; так, подошвенный Р. в связи с функцией ортоградности имеет у взрослого человека иную форму по сравнению с той, к-рая наблюдается в детском возрасте до выработки функции хождения в вертикальном положении. Далее, нек-рые* из двигательных актов, имеющие в детском возрасте чисто рефлекторный, автоматический характер, оказываются в дальнейшем под контролем коры, т. е. стимулируются или тормозятся корой, и т. о. превращаются из рефлекторного акта в волевой: примером этому может служить акт мочеиспускания, к-рый представляет собой рефлекторное явление у ребенка и волевой акт у взрослых. Анатсмо-физиологическаястру-к т у р а рефлекторного акта отчасти определена данными, приводившимися выше. Как уже указывалось, в осуществлении Р. принимают участие два или несколько невронов, находящихся между собой в синаптическом контакте. Совокупность этих невронов обозначается термином рефлекторная дуга. Соответственно основной сущности рефлекторного акта—восприятие внешнего раздражения и приведение к двигательному аппарату—в рефлекторной дуге имеются два основных элемента (неврона): приводящий (рецепторный, афферентный) и отводящий (эффекторный, эфферентный). Поэтому наиболее простой схемой рефлекторной дуги. являются два неврона—один, воспринимающий. внешнее раздражение и проводящий соответствующий этому раздражению импульс, до эф-фекторного неврона, по к-рому этот импульс проводится далее к периферическому двигательному аппарату, т.е. к мышце. В простейшем случае рефлекторная дуга представлена двумя; невронами: рецепторным и эффективным (рис. 1
А). Однако такая форма Р., осуществляемого» всего двумя невронами, представляет собой редкость; обычно рефлекторные дуги состоят из-нескольких невронов («цепь невронов»). В подавляющем большинстве случаев распространение импульса с рецепторного неврона на эффекторный происходит не непосредственно, между ними имеется т. н. вставочный или коррелятивный неврон (рис. 1
В). Фнкц. роль коррелятивного неврона состоит в том, что при помощи. его данный рецепторный неврон приходит в контакт с различными, иногда весьма отдаленными эффектор ными невронами (рис. 1 D, а, также рис. 2). Далее, данный рецептор при помощи ряда вставочных невронов может соединяться с одним эффектором (рис. 1
Е), благодаря чему происходит усиление, интенсивности импульса, т. к. каждая клетка'представляет собой. как бы резервуар скрытой энергии; под влиянием приносимого импульсом раздражения эта скрытая энергия переходит в кинетическую; т. о_ прохождение импульса через ряд вставочных, невронов может усиливать его интенсивность.. Возможны и другие более сложные сочетания: рефлекторных механизмов и соответственно-этому более сложные рефлекторные функции. Так, два рецепторных неврона могут сочетаться при помощи двух вставочных невронов с двумя различными группами эффекторных невронов-(рисунок 1
F). В этом случае раздражение каждого из двух рецепторов может вызвать реакцию обоих эффекторов. Если оба вставочных неврона оказывают стимулирующее влияние на один эффектор (или группу их) и тормозящее-влияние на другой эффектор (или группу их),. то это приведет к реакции в одной группе мышц при торможении в другой. Физиол, значение стимулирования одной группы мускулатуры при торможении другой заключается в том, что-благодаря этому может быть получен максимальный эффект рефлекторного движения при малой затрате энергии. Такой способ иннервации представляется чрезвычайно распространенным; он лежит в основе т. н. реципрокнои. («взаимно-противоположной») иннервации, т. е. .расслаблении антагонистов рефлекторно сокращающейся мускулатуры. Т. о. между системой рецепторных и эффекторных невронов заложен чрезвычайно сложный механизм сочетательных невронов. Следствием такого соотношения между рецептор-ными и эффекторными невронами является тот-факт, что каждый эффекторный неврон (т. е. периферический двигательный неврон) является общим проводником для импульсов, возникающих в различных отделах рецептивной поверхности; другими словами, один и тот же эффекторный аппарат может находиться под. воздействием различных рецепторов и в этом смысле представляет собой общий конечный путь-или последний общий этап (final common path). Выбор конечного пути, т. ё. направления, возникшего в данной рецепторной области раздражения в тот или другой периферический аппарат, определяется рядом различных условий: интенсивностью раздражения, его природой, локализацией его воздействия, предшествующими раздражениями, состоянием мускулатуры и др. Для иллюстрации влияния последних факторов могут быть приведены следующие приме-'ры. Если у обезьяны раздражать центральные концы Cvn и CVІii, то это раздражение может вызвать как сгибание, так и разгибание в локтевом суставе: если раздражению этих нервов предшествовало раздражение Di, получается разгибание; в том же случае, когда ему лредшествовало раздражение CVІ, получается сгибание (Sherrington). Что касается значения •состояния мускулатуры, то в этом отношении •следует упомянуть о законе Икскюля (Uex-ktill)* согласно к-рому возбуждение всегда распространяется к мышце, находящейся в состоянии растяжения, т. е. при возможности направления импульса в разные «конечные общие пути» •он, при прочих равных условиях, направляется в конечный путь, т. е. в периферический неврон, к-рый соответствует мышце, находящейся в «состоянии растяжения. Этот закон, установленный экспериментально, не вполне оправдывается клин, явлениями. Принципу конечного общего пути принадлежит важная роль сочетаний действия различных рефлекторных дуг при образовании рефлекторного акта. Последний никогда не представляет •собой результата действия одной рефлекторной .дуги, как это принималось нами выше для упрощения анализа рефлекторного механизма. Равным образом и раздражения обычно не бывают одиночными, как это принимается для простоты в приведенных выше схемах. На самом деле жаждый Р. представляет собой результат сложного взаимодействия сочетательных функций и участия нескольких рефлекторных дуг. Окончательная форма рефлекторного движения определяется теми конечными путями, по к-рым, в зависимости от упоминавшихся выше факторов, направляются импульсы. Взаимоотношение между рефлекторными реакциями может быть различным: нек-рые из них гармоничны, взаимно содействуют друг другу; такие Р. обозначаются термином аллиированные Р. Напротив, другие Р. представляются несовместимыми (напр. сгибание и разгибание); они не только не содействуют, но противодействуют друг другу. Такие Р. обозначаются термином антагонистические?.; антагонистические Р. оказывают друг на друга тормозящее влияние; тот Р. или группа Р., к-рые при этом взаимном торможении берут перевес, обозначаются термином препотентный (пересиливающий) Р. Аллиированные и антагонистические Р. представляют собой примеры суммацииР. Эти явления суммации и определения конечного пути составляют, как это предполагается многими, функцию синапсов. К явлениям суммации относится также индукция: взаимное усиление двух раздражений, воздействующих
1"последовательно или совместно (симультанная индукция) на две различные точки одного и того же рецептивного поля или рефлекторной зоны, т. е. области, из к-рой может быть вызван данный Р. Так например два воздействия, лежащие ниже порога раздражения, т. е. недостаточные каждое в отдельности для получения известного двигательного эффекта, вызывают этот последний, если они прилагаются совместно или последовательно в двух различных точках соответствующего рецептивного поля. Примером суммации последовательно действующих раздражений может служить прием штрихового раздражения, к-рым пользуются в клинике при исследовании кожных Р.; в то время как однократным прикосновением концом рукоятки молоточка нельзя вызвать подошвенного или брюшного Р., последние легко могут быть вызваны при проведении тем же предметом на известном протяжении по кожной поверхности. Координирующая и определяющая «конечный путь» роль синапсов центральной нервной системы лежит также в основе явления, обозначаемого термином центральное отражение. Под этим термином разумеется возможность превращений в форме и интенсивности Р., к-рые обусловливаются различными состояниями центрального аппарата. Для классификации Р. можно исходить из различных критериев. Соответственно месту возникновения возбуждения, то есть соответственно тем видам рецепторов, на которые действует раздражение, можно различать следующие формы Р.:экстероцептивныеР., обусловленные раздражением поверхностных (кожных) рецепторов или т. н. «контактцепто-ров»; при этой форме раздражение вызывается непосредственным соприкосновением поверхности тела с каким-либо предметом. Интеро-цептивнымиР. называются такие, в основе к-рых лежит раздражение «внутренней поверхности» тела, т. е. поверхности пищеварительного тракта, дыхательных органов и т. п. Т е -лерецептивными называются Р., при к-рых раздражение падает на т. н. дистансрецеп-торы, воспринимающие внешнее раздражение на расстоянии (зрительный, слуховой, обонятельный рецепторы). Наконец 4-ю группу данной классификации составляют т.н.проприо-цептивныеР., обусловленные раздражениями глубоких рецепторов, заложенных в мышцах, связках, суставах. К этой же группе могут быть относимы и рефлекторные импульсы, исходящие из вестибулярного аппарата. Физиол. роль проприоцептивных Р. состоит в координации движений, регулировании статики и положения тела. С точки зрения биологической Р. представляют собой приспособление двигательных реакций к действующим на организм влияниям внешней среды; в этом смысле можно говорить об известной фнкц. роли или биол. «назначении» Р. На основании этого критерия Р. могут быть подразделены на две группы: протективные, или защитные Р., устраняющие организм в целом или отдельные его части от вредных разрушительных внешних воздействий, и Р., воспроизводящие элементарные движения, входящие в состав определенного двигательного акта; эти последние Р. содействуют осуществлению функции известного органа или сами по себе представляют элементарный двигательный акт и имеют характер физиол. целесообразности и двигательной инициативы. — Относящиеся к первой группе протективные, или защитные Р. обозначаются также термином ноцицептив-ные Р. Ноцицептивные Р. отличаются следующими характерными особенностями. В противоположность остальным Р., для осуществления к-рых необходимы б. или м. определенные, специфические («адекватные») формы раздражений, ноцицептивные Р. могут вызываться любым раздражением, если оно достигает известной интенсивности и угрожает повреждением. Ноцицептивные Р. сопровождаются специфическим аффективным тоном неприятного переживания и имеют своим психическим коррелятом ч у в с т - во боли. НоцицептивныеР. препотентны,т. е. при возможности различных Р. на данное раздражение берет перевес ноцицептивная реакция, т. е. импульсы направляются по тому конечному пути, к-рый приводит к движениям, имеющим целью устранение всего организма или отдельной его части от внешнего воздействия. Эта препотентность придает ноцицептив-ным рефлекторным движениям характер непреодолимости, насильственности. Одним из типичных примеров ноцицептивных реакций может служить корнеальный Р. Что касается второй группы Р., носящих характер целесообразных двигательных актов не защитного характера, они представляют чрезвычайное разнообразие в отношении их формы и физиол. значения в смысле их большей или меньшей фнкц. оформленно-сти. В общем эти Р., обозначаемые термином «содействующие» (т. е. участвующие в волевых двигательных актах—fordernde Reflexe), включают в себя всю совокупность рефлекторных реакций за исключением ноцицептивных. К этой группе принадлежат разнообразные рефлекторные движения, начиная от изолированных мышечных сокращений и кончая сложными комплексами их, носящими черты целесообразности и относительной законченности в форме двигательных актов. В этом отношении необходимо иметь в виду следующее. Как указывалось выше, эволюция нервной системы представляет собой постепенную надстройку новых фнкц. элементов над старыми; при этом надстраивающиеся, филогенетически более новые корковые элементы подчиняют себе более старые сегментарные (спинальные) механизмы. Поэтому автономность спинальных Р. у человека представляется мало выраженной; Р. в узком смысле этого слова (см. выше), т. е. автоматические двигательные реакции спинного мозга, представляют собой у человека как бы аморфный материал, регулируя к-рый кора головного мозга формирует сложные двигательные акты, называемые волевыми движениями. Напротив, у низших животных с несовершенно развитым головным мозгом спинной мозг обладает значительной автономностью; совершаемые этими животными движения в значительной степени представляют собой рефлекторные автоматические акты, осуществляемые собственным аппаратом спинного мозга. Итак, у высших млекопитающих и особенно у человека рефлекторные автоматизмы находятся в б. или м. лятентном состоянии в виду подчинения их корковым функциям. Изучение этих чисто спинальных рефлекторных реакций возможно при условиях разобщения спинного мозга с головным, что достигается у животных экспериментально перерезкой спинного мозга в верхних отделах. В этом отношении заслуживают интереса исследования филипсона (Philippson) и Шеррингтона; исследования эти с несомненностью доказывают существование спииального рефлекторного механизма локомоции, т. е. актов ходьбы, бега и др. Эксперименты Филипсона состояли в отделении у собак поясничного отдела спинного мозга от вышележащих отделов центральной нервной системы. При этом оказалось, что у такого животного, подвешенного в воздухе, путем раздражения подошвенной поверхности одной из задних конечностей можно вызвать весь цикл мышечных сокращений, вполне воспроизводящих движения, свойственные задним конечностям при ходьбе. В этом случае раздражение одной конечности является исходным моментом всего цикла локомоторных движений: рефлекторное сгибание этой конечности является источником проприоцептивных Р. в форме сгибательных и раз-гибательных сокращений, требующихся для акта ходьбы. Так. обр. в сегментах поясничного отдела спинного мозга имеется интрасегментарный механизм локомоторной координации движений. Опыты Шеррингтона состояли в перерезке спинного мозга в более высоком отделе, над сегментами, соответствующими передним конечностям; т.о. отделенными от головного мозга оказывались спинномозговые сегменты, соответствующие как передним, так и задним конечностям. Раздражение одной передней конечности у такого «спииального» животного^ являлось стимулом к егибательным и разгибательным сокращениям всех четырех конечностей в той последовательности, к-ра^? имеет место при акте ходьбы. Эти эксперименты показывают, что рефлекторный механизм локомоторной координации движений представлен не только интрасегментарно и интерсегментарно, т. е. он может осуществляться не только в пределах сегментов, соответствующих задним или передним конечностям, но при помощи связей между шейными и поясничными сегментами может воспроизводить функцию ходьбы всеми четырьмя конечностями. Варьируя различным образом интенсивность раздражений, Шеррингтон мог получать у «спииального» животного и отдельные составные элементы акта ходьбы, как «Р. переступа-ния» (stepping-reflex), P. выбрасывания вперед передней конечности и др. Далее, опыты Шеррингтона показывают, что помимо локомоторных механизмов в спинном мозгу имеются рефлекторные механизмы других сложных актов (чесание, сотрясение тела и т. п.). У человека явления-спинального автоматизма наблюдаются при симптомокомплексе поперечного поражения спинного мозга; как уже указывалось выше, у человека в виду значительного развития коры автономность спинного мозга представляется мало выраженной; рефлекторная координация. локомоторных движений проявляется при этом лишь в рудиментарной форме, именно в виде т. н. «защитного Р.» или «реакции укорочения и удлинения»: раздражение одной из нижних конечностей вызывает ее сокращение в тазобедренном, коленном и голеностопном суставах, а вместе с этим разгибание во всех суставах противоположрой конечности; т. о. на стороне раздражения получается укорочение конечности, а на противоположной стороне удлинение,, и раздражая то одну то другую конечность, можно наблюдать попеременные сгибания одной конечности при разгибании другой, т. е. ту последовательность двигательных явлений, которая наблюдается в нижних конечностях при ходьбе. Существованием этого автоматического спинномозгового механизма объясняется возможность акта ходьбы без специального обращения внимания на совершаемые при этом движения. В нормальных условиях спинальные механизмы локомоторных актов находятся под регулирующим влиянием вышележащих отделов центральной нервной системы. Кора головного мозга оказывает на них тормозящее влияние, т. к. автономная деятельность спинного мозга исключала бы возможность осуществления корковых функций, т. е. так наз. волевых или сознательных двигательных актов. Важная роль в автоматическом регулировании спинальных механизмов принадлежит среднему мозгу. Как показывает новейшее исследование (см.
Магнус-Клейна рефлексы и
Посту-ральные рефлексы), эта регуляция имеет место не только при движениях, но и в покое; различного рода состояния покоя не представляют собой отсутствия иннервации, как это допускалось раньше, но имеют в своей основе специальные иннервации, источником к-рых являются mesencephalon и некоторые отделы стволовой части головного мозга (система вестибулярного нерва). «Спинальное» животное, т. е. животное с перерезкой спинного мозга в верхнем его от- деле, может, как указывалось выше, обнаруживать различные рефлекторные движения, но такое животное не может стоять вследствие отсутствия постурального тонуса в мускулатуре. Постуральный тонус в форме «Р. стояния» характерен для т. н. децеребрационной ригидности (см.
Децеребрация, децеребрационная ригидность). Описанные постуральные Р. обозначаются также термином «миостатические рефлексы». Помимо приведенных групп Р. заслуживают упоминания следующие. — Координированными Р, называются автоматические двигательные реакции, требующие для своего осуществления различных, иногда весьма удаленных друг от друга мышц и стало быть сегментарных центров спинного мозга. Примером координированных Р. может служить кашле-вой Р., в осуществлении которого принимают участие брюшные мышцы, диафрагма, межреберные мышцы, мускулатура гортани и другие (рисунок 2). Далее, примером коорди- нированного Р. может служить рвотный Р. (см.
Рвота).—О б щ и м и Р. называются двигательные рефлекторные реакции со- стороны всех четырех конечностей, если они развиваются одновременно. Примером такого общего Р. могут служить упоминавшиеся выше Р. при децеребрационной ригидности; поэтому они обозначаются иногда -также термином «общие статические Р.». Сюда же относится рефлекс Мора, или т. н. Р. обнимания; он наблюдается в раннем младенческом возрасте (первая неделя) и состоит в следующем: при различного рода раздражениях (сотрясения поверхности, на которой лежит ребенок, постукивание по животу, дуновение в лицо и др.) у ребенка наблюдается сначала разгибание и отведение всех четырех конечностей и тотчас же вслед за этим их сгибание и приближение к туловищу. Термином психорефлекс обозначаются рефлекторные реакции на эмотивные переживания; психорефлексы наблюдаются гл. обр. в области симпат. иннервации (сердце, сосуды, потоотделение и пр.). Как показывает явление т. н. «психогальванического Р.», рефлекторные реакции наблюдаются и при более тонких псих. процессах; психогальванический . рефлекторный феномен состоит в обнаруживаемых гальванометром электрических изменениях в коже при таких психических процессах, как воспоминание об аффекте, напряжении внимания и т. п. Ближайшая физиол. основа психогальванического Р. в точности не установлена (изменения влажности кожи, изменения в сосудах). Во всяком случае психогальванический Р.„ подобно рефлекторным изменениям дыхания и пульса при псих, процессах, должен быть поставлен в зависимость от симпат. иннервации.. Делались попытки установления Р. на псих. процессы со стороны произвольной мускулатуры, для чего пользовались так наз. «мультипликатором Зомера» — аппаратом, предназначенным для регистрации тончайших сокращений мускулатуры. Чрезвычайно важная роль принадлежит Р. в клин, исследовании нервной системы. С точки зрения симптоматологического их значения Р. могут быть подразделены на две главные группы: 1) Р. в е г е т а т и вной нервной системы, т. е. рефлекторные явления, имеющие-своим эффекторным аппаратом гладкую мускулатуру, и 2) Р. соматической нервной системы, имеющие своим эффекторным аппаратом поперечнополосатую мускулатуру. Р. соматической нервной системы классифицируются по месту возникновения возбуждения: различают глубокиеР., вызываемые раздражениями, к-рые наносятся на глубокие ткани (сухожилие, надкостницу), и поверхностные, вызываемые раздражениями кожи и слизистых оболочек. Техника вызывания глубоких Р. состоит в нанесении удара перкуторным молоточком по соответствующему сухожилию или надкостнице; кожные' Р. вызываются обычно штриховым раздражением кожной поверхности, а слизистые—прикосновением к слизистой оболочке или проведением по ней каким-либо предметом. Рефлексогенная зона Р. представляется довольно обширной, не ограничиваясь.
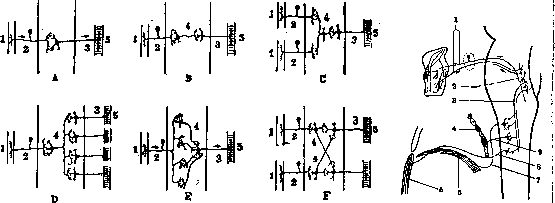
Рисунок 1. Рисунок 2.
Рисунок 1. Схема соотношения невронов при различных формах рефлекторного механизма:
J—рефлексогенная зона; 2—рецептор;
3—эффектор;
4—сочетательный (коррелятивный) неврон; 5—мышца. Рисунок 2. Схема кашлевого рефлекза. Импульс вследствие раздражения слизистой гортани по чувствительным волокнам X нерза распространяется в чувствительное ядро, а отсюда на двигательные волокна X нерва, а также при посредстве вставочного неврона (tr. solit'ario-spinalis) на спинномозговые центры, соответствующие диафрагме, межреберный и орюшным мышцам: 2—двигатзльные и чувствительные волокна (X нерв); 2—двигательные и чувствигельныз ядра X нерва;
3—сочетательный неврон между чувствительным ядром X нерва и спинным мозгом (tr. solitario-spinalis);
4—межреберные мышцы;
5—брюшные мышцы;6—диафрагма;
7—п. phrenicus;
8—нерв к мышцам живота;
9—нерв к межреберным мышцам. каким-либо одним нервом или сегментом; рефлекторные дуги большинства Р. соответствуют II—III спинномозговым сегментам; кроме того распространенность рефлексогенной зоны подлежит нек-рым индивидуальным колебаниям; она изменяется также в пат. состояниях, увеличиваясь при повышениях Р. и уменьшаясь мри их понижении. В прилагаемой таблице приводятся наиболее важные клин. Р. с указанием приемов их вызывания и уровня прохождения соответствующих им рефлекторных дуг через спинной мозг. Патологические изменения Р. соматической мускулатуры могут быть количественными (изменения интенсивности) и качественными (изменения формы). Что касается количественных изменений, то следует иметь в виду, что интенсивность Р. подлежит нек-рым индивидуальным колебаниям в физиол. условиях. Поэтому абсолютная интенсивность Р. сама по себе не имеет существенного значения для диагноза органического заболевания нервной системы при условии полной равномерности в интенсивности Р. с обеих сторон. Лишь крайние степени понижения или повышения Р. могут служить основанием для подозрения на органическое заболевание. Напротив, неравномерность сухожильных или кожных Р. всегда является указанием на наличие органического заболевания, природа и локализация к-рого могут быть установлены на основании сопоставления неравномерности Р. с другими симптомами.—Для графического воспроизведения рефлекторного акта и цифрового •определения интенсивности Р. предлагались различные приборы: рефлексограф, или рефлексометр В е й л е р а предназначен для записи на кимографе кривой коленного Р., точнее—изменения угла между голенью и бедром при осуществлении коленного Р. Сущность аппарата состоит в перекинутой через блоковид-ные колеса нити, к-рая связана, с одной стороны, с пером кимографа, а с другой стороны— перекинута через колесо, соединенное осевым стержнем с повязкой, обхватывающей толщу голени. Удар по сухожилию четырехглавой мышцы производится особым молоточком, рукоятка к-рого своей средней частью прикреплена поперек цилиндра, вращающегося на оси. При нагибании конца рукоятки ударйый конец молоточка приподьщается, а при внезапном прекращении надавливания на рукоятку ударный конец опускается и производит удар по сухожилию; т. о. сила удара не зависит от произвола исследователя. Кроме того она может быть определяема точно в цифрах передвижением груза, двигающегося по рукоятке молоточка между его ударной частью и местом прикрепления молоточка к цилиндру. Аппарат рассчитан таким образом, что при изменении угла между бедром и голенью на 1° пишущее перо кимографа проводит линию в 1
мм. С помощью рефлексотрафа Вейлера можно определить размер рефлекторного движения, время течения рефлекторного акта и отдельных его частей; так, Вейлеру удалось установить время скрытого периода коленного Р. (0,045—0,05 сек.). Рефлексометр Бехтерева представляет собой прибор для цифрового определения размаха голени при коленном Р.; прибор состоит из дугообразной градуированной пластинки, укрепленной поперек вертикального штатива. Размах голени может быть отсчитан по делениям дуги, параллельно к-рой двигается стопа при вызывании коленного Р. Описанные аппараты, подобно многим другим, предлагавшимся с той же целью, не получили применения в клин. практике гл. образ, потому, что необходимые для клин, выводов данные о состоянии Р. могут быть получены без этих приборов, отличающихся сложностью, а часто и неточностью. Что касается зависимости количественных изменений Р. от различных пат. условий, то всякого рода нарушения проводимости рефлекторной дуги приводят к ослаблению (гипореф-лексия) или утрате Р. (см.
Арефлексия). Поэтому утрата или ослабление Р. являются важным признаком периферических параличей. При пара» личах, зависящих от поражения центрального двигательного неврона, изменения со стороны Р. представляются более сложными и разнообразными. Нек-рые из Р. оказываются при этом повышенными (гиперрефлексия) вследствие выпадения тормозящих влияний коры на рефлекторный аппарат спинного мозга; другие Р. оказываются ослабленными; это касается тех Р., к-рые не тормозятся, а стимулируются корой. Кроме этих количественных изменений Р. центральные параличи сопровождаются качественными изменениями Р.: для этого рода параличей характерно появление таких рефлекторных реакций, к-рые не представляют собой ни повышения ни понижения Р., а являются своеобразными реакциями, не свойственными нормативному взрослому человеку. Совокупность этих реакций обозначается термином патологические Р. Сущность количественных изменений Р. при центральных параличах формулируется обычно таким образом, что при этих параличах глубокие Р. повышаются, а кожные понижаются. Это положение не может считаться правилом: при резких степенях центрального паралича наблюдается т, н. «защитный Р.» на кожные раздражения; в области губной мускулатуры при центральных параличах (псевдобульбарный паралич) наблюдается совершенно параллельное повышение как кожных, так и глубоких Р. (см. ниже); в области брюшной мускулатуры при центральных параличах наблюдается параллельное ослабление как кожных, так и глубоких Р. Все эти факты показывают, что ослабление или повышение Р. при центральных параличах не может ставиться в зависимость от их принадлежности к группе кожных или глубоких Р. Фактором, определяющим количественные изменения Р. при центральных параличах, являются биол. причины. При поражении центрального двигательного неврона спинной мозг оказывается отделенным от коры головного мозга; вследствие этого восстанавливается автоматическая деятельность спинного мозга, тормозящаяся в норме корковыми влияниями; но вместе с этим выпадают или ослабляются все те реакции спинного мозга, которые вырабатываются под влиянием мозговой коры и которые ею стимулируются. Кора головного мозга есть филогенетически поздняя надстройка над спинным мозгом, и явления, наблюдающиеся при поражении центрального неврона, т.е. при изоляции спинного мозга от влияний мозговой коры,могут быть рассматриваемы как регресс на более старый стадий эволюции. В спинном мозгу, возвращающемся при этом к своему автономному состоянию, исчезают те реакции, которые стимулируются корой, и усиливаются те, которые свойственны его собственному механизму и к-рые тормозятся корой. Так. обр. если при Таблица рефлексов. Название Р. Способ обнаружения Двигательный эффект Уговень рефлекторной дуги Анальный Р. ! Введение пальца в rectum Сокращения сфинктера Sv ' Ахиллов р. Постукивание по ахиллову сухожилию Сокращение икроножных мышц Si и Sn j 3 4 Бедренный—Ремака (наблюдается в пат. случаях—поражение верх-i их отделов спинного мозга) Biceps, P. с двуглавой мышцы Штриховое раздражение передне-внутреннего отдела верхней трети бедра Сгибание первых трех пальцев и стопы к подошве, разгибание в коленном суставе Нижний поясничный и верхний кре-стдовый сегменты Постукивание по сухожилию двуглавой мышцы плеча Сгибание в локтевом суставе Cv-VІ Верхний брюшной (син. супраумбили- кальный) Штриховое раздражение кожи живота кверху от пупочной линии Смещение брюшной стенки в сторону раздражения DVІi-VІii Гипохондральный Штриховое раздражение в области V—VІ межреберных промежутков Сокращение верхнего отдела прямой мышцы Dv-VІ Глоточный Прикосновение к задней стенке глотки Кашлевое движение Чувствит. и двига-тельн. ядра IX нер. Infraspinatus, P. с надкостницы spinae scapulae Постукивание по лопатке книзу от spinae scapulae Приведение плеча, ротация кнаружи, сгибание в плечевом суставе Cv-VІ Коленный (син. пате-лярьый) Постукивание по сухожилию четырехглавой мышцы Разгибание в коленном суставе Ln-iv Кохлео-пальпебраль-ный (син. слуховой) Внезапный звук Смыкание век Ядра кохлеарного и лицевого нервов Лопаточно-плечевой (scapulo-humeralis) Постукивание по внутреннему краю лопатки Приведение плеча, ротация его кнаружи, иногда сгибание в плечевом суставе
i Civ-v Лопаточный, s.' spino- scapularis (син. скапу- лярный) Постукивание по внутреннему краю лопатки или штриховое поверхностное раздражение кнутрн от энутреннего края лопатки Приведение лопатки Cv-VІ
н Masseter-reflex (син. нижнечелюстной, ман-. дибулярный) При открытом (без напряжения) рте исследуемого производят короткое постукивание но резцам нижней челюсти Сокращение жевательных мышц Чувствит. и двигателей. ядра тройничного нерва Надбровный Постукивание по внутреннему краю надбровной дуги Смыкание века Ядра тройничного и лицевого*нервов Нёбный, Р. с uvulae Прикосновение к мягкому нёбу Приподымание uvulae Чувствит. и двига-тельн. ядра блуждающего нерва Нижний брюшной (син. инфраумбили- нальный) Раздражение кожи живота книзу от пупочной линии Смещение брюшной стенки в сторону раздражения Dx-xn Пальцевой (рефлекс Майера) Резкое сгибание среднего пальца к ладони, производимое исследователем Оппозиция большого пальца, сгибание его в пястно-фалан- говом и разгибание в межфа- ланговом сочленении Cvn-VІn и
Их Подошвенный Штриховое раздражение кожи подошвы Сгибание пальцев к подоите Si и Sn Предплечья (Лери) Сгибание с силой и одновременно пальцев и кисти руки Рефлекторные сгибания верхней конечности в локтевом суставе Cti Р. с кремастера, Р. т. cremasteris, P. с яичка Штриховое раздражение внутренней поверхности верхнего . отдела бедра Прин#дымание мошонка Li-п £
% •
Название Р. |
Способ обнаружения Двигательный эффект Уровень рефлекторной дуги P. m. tensoris fasciae latae (P. Brissaud) Штриховое раздражение кожи Сокращение m. tensoris fasciae подошвы j latae Si и Sn 22 23 24 25 P. с локтевой кости (син. улънарный, ку- битальный) Постукивание по шиловидно- | му отростку локтевой кости Пронация и сгибание в локтевом суставе CVІ Р. с лучевой кости (син. карпо-радйаль- ный Р.) с processus styloideus radii Постукивание по шиловидному отростку лучевой кости Сгибание в плечевом суетаве, пронация, сгибание пальцев Су ДО D
x Роговичный, Р. с cornea Прикосновение к роговой оболочке Смыкание век Ядра тройничного и лицевого нервов Сосательный (нормально имеется лишь в младенческом возрасте) Прикосновение к губам Присасывающие движения Ядра тройничного и лицевого нервов Стернальный Р. При раздражении кожи подложечной области Сокращение брюшных мышц 27 28 Triceps, P. с трехглавой мышцы Постукивание по сухожилию трехглавой мышцы плеча Разгибание в локтевом суставе CVІ-vn Феморальные (Guillain u. Barre): а) Тибио-фемораль- ный б) Перонео-фемораль-ный Постукивания по сухожилиям mm. semimembr. и semitend. (больной находится в лежачем положении на спине с несколько отведенной и слегка согнутой нижней конечностью) Постукивание по сухожилию (больной находится в лежачем положении на боку, противоположном стороне исследования) Сокращение mm. semimemhra-nosi и semitendinosi Сокращение m. biceps femoris Ljv-v Si
I 29 i Чихательный Щекотание слизистой оболочки носа Чиханье Ядра тройничного и блуждающего нервов j 30 Ягодичный (син. глютеальный) Штриховое раздражение кожи ягодичной области ■■ » Сокращение большой ягодичной мышцы i Liv-v и Si центральных параличах ослабляется или исчезает подошвенный Р., то это объясняется не тем, что этот
Р. кожный, а тем, что этот Р. филогенетически и онтогенетически новый: он отсутствует у ребенка и приобретается лишь в течение индивидуальной жизни в процессе приспособления стопы к функции хождения в вертикальном положении. Равным образом и брюшные Р. угасают или ослабляются не потому, что они кожные, а потому, что они приобретаются в течение индивидуальной жизни в связи с вертикальным положением тела; подобно подошвенным Р. брюшные Р. представляют собой реакцию, стимулируемую корой; они отсутствуют у животных, а также у детей до приобретения ими функции стояния и сидения. Наоборот, повышение коленного Р. при центральном параличе зависит не от его принадлежности к группе глубоких Р., а от того, что функция разгибания на уровне коленного сустава есть фи-. логенетически старая, свойственная всем млекопитающим и поэтому автоматизировавшаяся, врожденная форма рефлекторной реакции, не стимулируемая, а тормозящаяся корой головного мозга. Приведенные биол. факты следует иметь в виду для объяснения тех исключений и противоречий, к-рые существуют в отношении общераспространенного положения о том, что при центральных параличах повышаются глубокие Р. и ослабляются или исчезают кожные. К области количественных изменений Р. может быть отнесен также факт увеличения рефлексогенной зоны при повышении Р. и ограничение ее при ослаблении Р. Особенно важная роль в диагностике поражений центрального неврона принадлежит упоминавшимся выше пат. Р. Под этим термином следует разуметь рефлекторные реакции, не существующие у нормального взрослого человека и появляющиеся лишь при поражении центрального двигательного неврона. К этой характеристике пат. Р. можно было бы прибавить еще одну важную для объяснения их сущности черту: все эти Р. свойственны младенческому или детскому возрасту; это обстоятельство может служить поводом к объяснению пат. Р. теми биол. данными, к-рые были приведены выше по отношению к количественным изменениям Р. Наличие реакций идентичных пат. Р. в раннем детском возрасте, т. е. до полного развития коры„ исчезновение их у взрослого по мере формирования корковых функций и наконец восстановление их при поражении пирамидных путей,. т. е. при изоляции спинного мозга от корковых влияний,—все это свидетельствует о том, что* пат. Р. представляют собой автоматические реакции спинного мозга, находящиеся у взрослого нормального человека в лятентном состоянии под влиянием корковой деятельности. По своему биол. смыслу они являются рудиментами функций, представляясь филогенетически старыми, зафиксировались и сделались автоматичными. При выработке корковых функций эти врожденные автоматизмы переводятся в скрытое состояние и заменяются новыми формами реакций, соответствующими корковым функциям. Из отдельных форм пат. Р. заслуживают упоминания следующие. В области лицевой мускулатур ы при двусторонних поражениях центральных невронов (tr. eortico-bulba-ris) наблюдается наклонность к рефлекторному сокращению губной мускулатуры (m. orbicu-laris oris); совокупность различных способов вызывания этих реакций с губной мускулатуры может быть обозначена термином «оральные Р.». При постукивании по корню носа («назо-лаби-альныйР.» Аствацатурова, глабелярный Р. Моро), при прикосновении к губам (Fressreflex Ор-penheim), при постукивании по верхней губе (рефлекс Toulouse и Vurpas, s. r. buccalis или хоботковый), при постукивании по резцам верхней или нижней челюсти, при прикосновении к твердому нёбу наблюдается сокращение круглой мышцы рта, приводящее в резких случаях к хоботкообразному смыканию губ-, иногда к нему присоединяются жевательные и глотательные движения. По своему физиол. смыслу упомянутые оральные Р. представляют собой рудимент сосательных движений. Эта реакция сосательных движений представляет собой филогенетически весьма старую функцию, автоматизировавшуюся на пути эволюции; «сосательный Р.» в качестве врожденного «безусловного» Р. существует и у человека в, младенческом возрасте, но по мере развития коры и замены автоматических сегментарных реакций губной мускулатуры произвольными корковыми, сосательный Р. переходит в скрытое состояние. При поражении пирамидных путей (псевдобульбар-ный паралич) происходит регресс на более низкую ступень эволюции и сосательный Р. восстанавливается из своего скрытого состояния. Весьма большое число пат. реакций описано в области стопы и пальцев ноги. Основным типом их является наклонность к тыльной флексии большого пальца и к разведению остальных пальцев йоги при поражениях пирамидного пучка. Впервые эти симптомы были описаны Бабинским в форме тыльной флексии большого пальца (см.
Бабииского рефлекс) и разведения пальцев («симптом веера») при штриховом раздражении кожи подошвы. Аналогичный рефлексу Бабинского трокмоторный рефлекс вызывается постукиванием спинки стопы и области метатарзо-фаланговых суставов кнаружи от сухожилия длинного разгибателя большого пальца. Со времени первого описания Бабинским тыльной флексии большого пальца (1895 г.) при указанном сейчас способе раздражения описывались другие способы' получения того же феномена: резкое надавливание сверху вниз по задней поверхности tibiae (см.
Оппенгейма рефлекс), надавливание на толщу мышц задней поверхности голени (см.
Гордона признак, рефлекс, феномен), сдавливание ахиллова сухожилия (симптом Шефера), давление на подошву (симптом Треммера) и др. Следует отметить, что при разных степенях пирамидного поражения наклонность к тыльной флексии большого пальца бывает иногда выражена настолько резко, что это явление может быть вызываемо самыми разнообразными раздражителями. Нужно однако признать, что наиболее надежным приемом обнаружения этого симптома является классический способ, рекомендованный Бабинским; это—штриховое раздражение острым предметом наружного края стопы. В этой форме тыльная флексия большого пальца представляет самый ранний симптом развивающегося поражения центрального-двигательного неврона; вместе с тем он нередко-остается единственным признаком пирамидного, поражения после полного восстановления двигательной функции.—-Вторую группу пат. Р. со стороны стопы составляют явления рефлекторного сгибания II — V пальцев; наиболее целесообразными способами обнаружения этого симптома являются:
Мендель-Бехтерева рефлекс (см.) (сгибание II—V пальцев при постукивании по наружной части тыла стопы) и симптом Рос-солимо (сгибание II—V пальцев при коротком ударе по концу III пальца). По своему биол. смыслу пат. Р. стопы представляют собой рудимент хватательной функции. Как известно, черты хватательной функции выражены довольно ясно в младенческом возрасте. Хватательная функция нижней (задней) конечности свойственна также всем ближайшим филогенетическим предкам человека. У взрослого человека эти черты хватательной функции переходят в скрытое состояние под влиянием приспособления стопы к статической функции. При поражении пирамидного пучка тормозящее влияние коры выпадает, и находившиеся в скрытом состоянии черты хватательной функции восстанавливаются: появляется подвижность пальцев ноги, наклонность к сгибанию II—V пальцев и разгибанию большого, а также наклонность к -ротации стопы кнутри(см.Гадршб"ерга
рефлекс),— все эти явления свойственны стопе в младенческом возрасте. Из пат. Р. верхней конечности заслуживает упоминания т. н. «хватательный Р.»; Сущность этого Симптома заключаемся в том, что больные, неспособные вследствие паралича произвести произвольно хватательное движение, захватывают автоматически предметы, помещаемые им в руку, причем это рефлекторное захватывание не может быть произвольно прекращено больным. Хватательный Р. наблюдается при поражении лобной доли и т. о. не является признаком повреждения пирамидного пучка; он представляет собой автоматизировавшуюся функцию двигательной корковой области (передняя центральная извилина); хватательный автоматизм существует, какизвестно, в младенческом возрасте, впоследствии он тормозится со стороны лобной доли, а при поражениях последней восстанавливается из скрытого состояния. Помимо указанного существуют и другие формы проявления хватательного Р.: так напр. иногда б-ные, страдающие поражением лобной доли, обнаруживают непроизвольное стремление захватить больной рукой предмет, находящийся в здоровой руке. Легкую степень хватательного Р. представляет собой т. н. «тоническая иннервация»; явление это состоит в невозможности произвольно прекратить хватательное движение при сохранности произвольного акта хватания; б-ной может захвастать предмет, но не в состоянии выпустить его из руки произвольно. В числе пат. изменений Р. следует упомянуть от. н. парадок сальных Р. Сущность обозначаемого этим термином явления состоит в том, что в нек-рых пат. случаях попытка вызвать определенный Р. приводит к рефлекторной реакции со стороны антагонистов тех мышц, на рефлексогенную зону к-рых было направлено раздражение; напр. при нанесении раздражения на сухожилия сгибателей получается рефлекторное сокращение разгибателей. Парадоксальные Р. могут наблюдаться как при центральных, так и при периферических параличах, При центральных параличах парадоксальные Р. обусловливаются неравномерностью повышения рефлекторной возбудимости в различных мышечных группах. Так например при спастических параличах рефлекторная возбудимость разгибателей коленного сустава повышается значительно больше, чем сгибателей, а вместе с этим чрезвычайно увеличивается и рефлексогенная зона. Это преобладание повышения возбудимости разгибателей по сравнению со сгибателями, а равно и распространение рефлексогенной зоны разгибателей может быть настолько значительным, что раздражение сухожилий сгибателей имеет результатом рефлекторное сокращение разгибателей. Парадоксальные Р. при периферических параличах наблюдаются редко, и в отношении объяснения их механизма существуют нек-рые разногласия. Следует впрочем допустить, что основной причиной парадоксальных Р. и в этом случае является иррадиация раздражения в несоответствующий сегмент вследствие резкого нарушения равновесия в рефлекторной возбудимости двух рефлекторных центров: более возбудимый центр реагирует при этом на раздражения, наносимые на рецепторы менее возбудимого.— Рефлекторный тонус, т. е. степень напряжения мускулатуры в покое, степень ее предуготованности к сокращению уже в физиол. условиях, представляется неодинаковой в различных мышечных группах; в пат. случаях эта разница может достигать еще более резких степеней. Р. вегетативной нервной систе-м ы не приобрели и повидимому не могут приобрести той степени диагностической точности и объективности, которая свойственна соматическим Р. Факт этот объясняется многими причинами: во-первых рефлекторные дуги вегетативной системы не представляют такой изолированности и автономности, какая свойственна соматической нервной системе; поэтому вегетативные Р. представляются более диффузными и распространенными; во-вторых рефлекторная возбудимость вегетативной нервной системы подлежит весьма значительным индивидуальным колебаниям; в-третьих весь аппарат вегетативной нервной системы представляет собой весьма неустойчивый механизм, равновесие к-рого может резко колебаться в физиол. условиях: такие моменты, как психические переживания, состояние органов пищеварения, утомления или физ. бодрости и целый ряд других обстоятельств, могут влиять на вегетативные Р. Наконец нельзя игнорировать и того, что изучение вегетативной нервной системы составляет достояние недавнего времени. Рецепторными аппаратами рефлекторных вегетативных дуг являются во-первых афферентные волокна симпатической нервной системы, а во-вторых афферентные волокна соматической нервной системы. Далее следует иметь в виду, что все соматические нервы содержат в своем составе и симпатические волокна. Поэтому источником вегетативных Р. могут быть раздражения, весьма разнообразные как в отношении своей природы, так и места их воздействия. Так например болевые раздражения, действующие на кожу, могут вызывать различные реакции не только со стороны вегетативных эффекторов кожи, но и со стороны внутренних органов в виду обилия анат.-физиол. связей между соматическими афферентными волокнами и симпат. центрами. — Систематической классификации вегетативных Р., основанной на каком-либо определенном принципе, не установлено, и различные виды вегетативных Р. группируются то по вызывающим их раздражителям (напр. «фармакологические Р.»), по тому органу, с к-рого получается реакция (например «вазомо-торрефлексы»), то по характеру этих реакций (напр. преесорные и депрессорные Р. и т. п.). Помимо описанных (см.
Вегетативная нервная система, афферентные пути и рефлексы вегетативной нервной системы, а также висцеро-рефлексы) вегетативных Р. упомянем о следующих их формах. К о ж н о-с ердечные Р.—изменения пульса в форме замедления или учащения при различных кожных раздражениях: болевых, осязательных, фарадических и др. Нёбно-сердечный Р.—ускорение пульса при давлении на твердое нёбо. Ортостати-ч е с к и е P. (Prevel)—учащение пульса при переходе из лежачего или сидячего положения в стоячее. О р т о с е р д е ч н ы й Р., наблюдающийся лишь в пат. случаях,—расширение сердечной тупости при переходе из горизонтального положения в вертикальное. Симптом Эрбен а—замедление пульса (раздражение п. vagi) при положении исследуемого на корточках с головой, резко нагнутой кпереди. Рефлекс Чермака—замедление пульса и дыхания при давлении на верхнюю треть гру-дино-ключично-сосковой мышцы. Брюшино-сердечный Р.—увеличение сердечной тупости при постукивании по средней линии живота между мечевидным отростком и пупком. К о х -леарный Р. на сердце—слуховые раздражения имеют своим результатом соматические Р. поворота глаз и мигания (см. табл.—кохлео-пальпебральный Р.); при более сильных раздражениях распространение импульса на ядро блуждающего нерва может вести к изменениям ритма дыхания и пульса. Р. солнечного сплетения (син. солярный Р.)—давление по средней линии живота до ^ощущения пульсации брюшной аорты кверху от пупка—замедление пульса. Общим термином преесорные Р. обозначаются явления повышения кровяного давления и сужения под влиянием различных воздействий. Обратные изменения обозначаются термином депрессорные рефлексы Мамидло (со-сково)-ареолярный Р.—сморщивание грудного соска при кожных раздражениях. Яичко-сердечный Р.—прижатие яичка—вызывает целый ряд явлений со стороны сердца, сосудов, дыхательных путей, зрачков. См. также
Аддукторный рефлекс, Антагонистические рефлексы, Ахиллов рефлекс, Ашнера феномен, Барре рефлекс, симптом, Беттигера рефлекс, Бехтерева рефлексы, Брюшные рефлексы, Бульбарныё рефлексы, Бульбо-кавернозный рефлекс, Вартенбер-га рефлекс, Водака рефлекс, Гнуди рефлекс, Губной рефлекс, Глабелярный рефлекс, Гирш-берга рефлекс, Гааба рефлекс, Геринга рефлекс, Даниелополу рефлексы, Жуковского рефлекс, Зрачковые волокна, рефлексы, центры, Кожные рефлексы, Контралятеральные рефлексы. Кор-неальный рефлекс, Конъюнктивалъиый рефлекс, Коленный рефлекс, Кремастера рефлекс, Магнус-Клейна рефлексы,, Мари-Фуа рефлексы, Надкостничные рефлексы, Пиотров&козо рефлекс, Пиломоторные волокна, рефлекс, Постуральные рефлексы, Россолимо рефлекс, Сухожильные рефлексы, Условные рефлексы. Лит.—см. лит. к ст.
Рефлексология. М. Аствацатуров.
РЕФЛЕКТОР, прибор для отражения в определенном направлении световых лучей, идущих из какого-либо источника. С целью наилучшего использования света для изготовления Р. пользуются или полированной поверхностью одного из светлых металлов—серебра, никеля—или посеребренной с внутренней стороны стеклянной поверхностью. Смотря по назначению, эта поверхность может быть плоской или вогнутой, причем в последнем случае кривизна может варьировать в весьма разнообразных формах: сферическая, параболическая и пр. В медицине употребление Р. сводится к освещению того или другого исследуемого участка и т. к. в каждой отдельной области медицины имеет место свое специальное применение Р., то они являются необходимой принадлежностью в общем наборе того или другого инструментария, иногда со специальной номенклатурой, как например офтальмоскоп (см.). См. также Ото-рино-лярин-гологический инструментарий, Ларингоскопия, Бронхоскопия, Зеркала, Хирургический инструментарий.
 Смотрите также:
Смотрите также:
- REFLUX, обратный ток, затекание жидкости (мочи или контрастного вещества) в противоестественном направлении; В урологии термин R. применяется 1) в отношении затекания мочи из мочевого пузыря в мочеточник и лоханку—пу-зырно-мочеточниковый R., ...
- РЕФРАКТЕРНАЯ ФАЗА, состояние временной невозбудимости, наблюдаемое в тканях после каждого одиночного имцульса возбуждения. Впервые Р. ф. найдена Мареем (Магеу, 1876) при изучении деятельности сердца; электрическое раздражение, наносимое сердцу в момент его ...
- РЕФРАКТОМЕТРИЯ (от лат. refringo—■ преломляю), метод измерения преломления (см.) светового луча, идущего .из одной среды в другую. При определении показателя преломления рефрактометром,^ е. i-воздух прибором, изме-------4 ряющим преломление, в качестве первой ...
- РЕФРАКЦИЯ (от лат. refringere — отражать, разламывать), оптическая установка глаза в. состоянии покоя аккомодации. Глаз как оптический аппарат построен по типу фотографической камеры. В глазу, так же как в фотографической камере, ...
- РЕЦЕПТ (от лат. receptum—взятое, от глагола recipere—брать обратно, взять), письменное предписание врача (praescriptio medica), до-| кумент, по к-рому а) врач выписывает лекар-I ство из аптеки для б-ного, ...