МЫШЕЧНАЯ СИСТЕМА
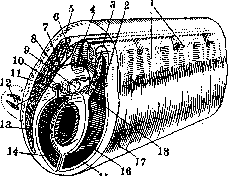
Рисунок 1. Поперечный разрез эмбриона позвоночных (схематично): 1— первичный сегмент; 2—dermatom; 3—myotom; 4—спинной мозг; 5— corium; 6—гангляон; 7—ramus dorsalis; 8 -ramus ventralis; 9—chorda; 10—позвонок; 11— ношка первичного сегмента; 12—конечность; 13—брюшной отросток; 14—полость тела; 15 — висцеральная мезодерма; 16—париетальная мезодерма; 17—эктодерма; 18—scle-rotom.
ных волокон, охватывающих весь висцеральный аппарат снизу и с боков и образующих общий его сжиматель (m. constrictor superficialis), а также из отдельных более глубоких мышц, прикрепляющихся к отдельным висцеральным дугам и приводящих их в движение (mm. adductores arcuum, in-terarcuales etc.). Эти мышцы иннервируются в области челюстной дуги тройничным нервом, в области подъязычной—лицевым, в области первой жаберной дуги—языко-глоточ-ным и в области следующих я-саберных дуг— блуждающим нервом. Соответственно большому значению висцерального аппарата его висцеральная мускулатура получает в области головы преобладающее развитие, меж- ду тем как соматическая мускулатура здесь отступает на задний план. В связи со значительным развитием челюстей особенно сильно развивается мускулатура первых двух дуг. Мышцы челюстной дуги распадаются на мышцу, поднимающую нёбно-квадратный хрящ (m. levatorpalato-quadrati; n. V), межчелюстную мышцу (m. intermandibularis; п. V, VII) и мышцу, приводящую нижнюю челюсть (m. adductor mandibulae; n. V). Последняя дает у наземных позвоночных всю жевательную мускулатуру, именно—собственно жевательную (m. masseter; n. V), височную (ш. temporalis) и крыловидные мышцы (тт. pterygoidei). Из передней части межчелюстной мышцы развивается подборо-дочно-подъязычная мышца (m. mylo-hyoi-deus). Из мышцы подъязычной дуги у наземных позвоночных получает особое значение мышца, прикрепляющаяся к заднему углу нижней челюсти и опускающая ее (т. depressor mandibulae). У млекопитающих она несколько видоизменяется и дополняется еще частью межчелюстной мышцы (m. di-gastricus, seu mandibulae; n. V, VII). У наземных позвоночных сильно разрастается поверхностный сжиматель области подъязычной дуги, дающий мышцу, охватывающую у рептилий шею снизу и с боков (m. sphincter colli; п. VII). У млекопитающих от этой мышцы отщепляется поверхностный подкожный слой (platysma myoides). Эта мускулатура иннервируется согласно своему происхождению лицевым нервом, разрастается на всю область головы и дает начало сложной системе подкожной лицевой мускулатуры, в том числе и мимической мускулатуре обезьян и человека. Мышцы собственно жаберного аппарата, принадлежащие области языко-глоточного и блуждающего нервов, с утратой жаберного дыхания у наземных позвоночных редуцируются, но частью сохраняются в виде мышц подъязычного аппарата и особенно—гортанных мышц. К висцеральным мышцам относится по своему происхождению также и трапециевидная мышца (m. trapezius, s. cucullaris; n. X), вступающая в связь с плечевым поясом. У высших позвоночных иннервирующая эту мышцу задн. часть блуждающего нерва обособляется в виде самостоятельного добавочного нерва (п. accessorius). У млекопитающих от трапециевидной мышцы обособляется нижняя часть, к-рая получает название грудино-ключично-сосцевидной мышцы (m. sterno-eleido-mastoideus; n. XI).— Соматическая мускулатура позвоночных вся поперечнополосата, развивается за счет мускульного листка миотома (первичного сегмента) и иннервируется нервами, волокна которых выходят из брюшных рогов и ядер мозга в составе брюшных корешков спинномозговых нервов (и соответствующих им головных нервов III, IV,VIhXII пар). Она сохраняет у низших позвоночных в течение всей жизни свое сегментарное строение. Именно такое расчленение мускулатуры обеспечивает достаточную подвижность при плавании в воде при помощи боковых извивов тела, при к-рых отдельные миомеры сокращаются последовательно. С развитием черепа и органов чувств у позвоночных од- нако теряется подвижность переднего, головного отдела, и соматическая мускулатура здесь редуцируется. Сохраняется лишь незначительная ее часть в виде глазных мышц, развивающихся из передних трех головных миотомов, и подъязычн. мускулатуры, развивающейся из брюшных отростков задних головных миотомов. У всех позвоночных имеются четыре прямые мышцы и две косые мышцы глаза, отходящие: первые от дна глазницы с задней ее стороны, а вторые от передней стенки глазницы. Верхняя, внутренняя и нижняя прямые и нижняя косая мышцы развиваются из первого миотома головы и иннервируются глазодвигательным нервом, являющимся нервом этого сегмента. Верхняя косая мышца развивается из второго миотома и иннервируется блоковым нервом, а наружная прямая мышца развивается из третьего миотома и иннервируется отводящим нервом. У наземных позвоночных от последней мышцы обособляется более глубокая порция, идущая прямо к основанию глазного яблока и втягивающая последнее внутрь глазницы (m. retractor bulbi). У обезьян и у человека эта мышца редуцируется.—Подъязычная мускулатура сохраняет у низших позвоночных иногда еще свою сегментацию и представляет собой продольную мускульную массу, лежащую с брюшной стороны висцерального скелета и соединяющую брюшные концы последнего с плечевым поясом. У наземных позвоночных эта мускулатура распадается на задний участок, идущий от плечевого пояса и грудины до подъязычного аппарата (m. sterno-hyoideus и т. omo-hyoideus), и передний участок, идущий от подъязычного аппарата до подбородка (m.genio-hyoideus). От последней обособляется и собственная мускулатура языка (m. genio-glossus и т. hyo-glossus). Вся подъязычная мускулатура иннервируется комплексом затылочных и иногда передних спинномозговых нервов, к-рый только ■у высших позвоночных становится типичным головным нервом (п. hypoglossus). Мускулатура туловища состоит у низших позвоночных из ряда миомеров правой и левой стороны, к-рые образуют т. н. боковые мышцы. Последние разделены горизонтальной соединительнотканной перегородкой на спинную и брюшную мышцы. У амфибий спинная мускулатура состоит еще из ряда независимых миомеров, но у рептилий и выше стоящих позвоночных миомеры разбиваются на отдельные мускульные пучки, связывающие между собой отдельные части соседних позвонков (mm. in-terspinales, intertransversales, transverso-spi-nales, transverso-costales), частью же соединяются для образования более длинных мышц (m. longissimus dorsi). —Б рюшная мускулатура диференцируется уже у высших рыб и у амфибий на отделы и слои, различающиеся направлением волокон. Посередине брюха обособляется продольная мускульная полоса—прямая мышца брюха (т. rectus abdominis), к-рая иногда еще подразделяется на глубокую и поверхностную. В боковых стенках направление волокон меняется и притом различно на различной глубине: в поверхностных слоях они направ- ляются косо назад к брюху, в более глубоких слоях—косо назад к спине, а в самых глубоких слоях принимают приблизительно поперечное направление. Т. о. развиваются и обособляются друг от друга наружная и внутренняя косые мышцы (mm. obliqui ех-ternus et internus) и поперечная мышца (m. transversus abdominis). — В области грудной клетки у высших позвоночных брюшные мышцы разделяются на отдельные участки, лежащие между ребрами и образующие наружную и внутреннюю межреберные мышцы (mm. intercostales).B поясничной области им соответствует квадратная мышца (m. quadratus lumborum).—Впереди от ребер та же мускулатура является в виде лестничных мышц (mm. scaleni). У млекопитающих наружная и внутренняя межреберные мышцы дают начало еще и дальнейшим продуктам диференцировки: задним зубчатым мышцам (mm. serrati postici super, et infer.), являющимся специально дыхательными мышцами. Дыхательной мышцей соматического происхождения является также и диафрагма, развивающаяся только у млекопитающих в поперечной перегородке, непосредственно позади сердца. Мускулатура конечностей развивается в виде ряда эпителиальных мускульных почек, вырастающих от нижних концов миотомов. Мускульные почки делятся каждая на верхнюю и нижнюю вторичные почки, к-рые врастают в закладку конечности, распадаются через разрыхление связи между клетками на мезенхиму и дают сплошной слой миогенных клеток на спинной и брюшной стороне закладки. У рыб они дифе-ренцируются на мускульные пучки, соответствующие лучам скелета плавников. У наземных позвоночных зачаток мускулатуры диференцируется сложнее и кроме того разрастается в области плечевого пояса на спину ,и грудь, где образует «первичную» мускулатуру плечевого пояса. В передней конечности из спинного мускульного зачатка так. обр. развиваются спинные мышцы плечевого пояса (m. deltoideus scapulae впереди, т. е. краниальнб, m. dor-salis scapulae над лопаткой и m. latissimus dorsi позади, т. е. каудально) и мышцы-разгибатели конечности (m. anconaeus, s. quadriceps, тт. extensores). У млекопитающих, особенно у приматов, за счет последних развиваются еще мышцы, поворачивающие конечность ладонью вверх (mm. supinatores).— Из брюшного зачатка развиваются брюшные мышцы плечевого пояса (m. coraco-brachia-lis впереди, т. е. краниально, и m. pectoralis позади, т. е. каудально) и все сгибатели конечности (m. coraco-radialis, s. biceps, mm. flexores). У лазающих млекопитающих за счет последних развиваются еще мышцы, поворачивающие'конечность ладонью вниз (mm. pronatores). Кроме того непосредственно за счет миомеров боковой мышцы позднее развивается еще «вторичная» соматическая мускулатура пояса, состоящая из мышцы, поднимающей лопатку (m. levator scapulae), и передних зубчатых мышц (mm. serrati anti-ci). В связь с плечевым поясом вступает также еще одна висцеральная мышца—-трапециевидная (m. trapezius).—В задней ко- нечности нет вторичной мускулатуры пояса. Одна из мышц, прикрепляющихся к свободной конечности, имеет однако вторичное происхождение из хвостовых мышц: это —■ лежащая позади грушевидная мышца (m. piriformis, s. caudo-femoralis). Остальные мышцы развиваются из первичных мускульных зачатков: приводящие мышцы (mm. adductores), гребенчатая (m. pectine-us) и лобково - бедренная (m. pubo-femora-lis) впереди и изнутри, ягодичная (m. glu-taeus, s. ilio-femoralis) позади и снаружи. В пределах свободной конечности разгибатели (m. quadriceps femoris, mm. extensores) представляют результат диференцировки первичной спинной, а сгибатели (m. biceps femoris, тт. flexores)—первичной брюшной мускулатуры КОНечнОСТИ. и. Шмальгаузен. П. Мышцы и их вспомогательные аппараты. Различаются три вида мышечной ткани: поперечнополосатая (произвольная), гладкая (непроизвольная) и сердечная (см. Сердце). Гладкая встречается в коже и в стенках внутренних органов, из поперечнополосатой построены мышцы скелета; кроме того произвольные мышцы имеются в нек-рых других системах: гортань, начало и конец пищеварительного тракта, конец мочеиспускательного канала, мышцы глаза и среднего уха. Поперечнополосатая мускулатура скелета представляет мягкую, ясно волокнистую, красно-бурого цвета массу, к-рая покрывает все части скелета, т. ч. свободными остаются только немногие выступы костей, легко поэтому прощупываемые через кожу. Мускулатура в целом у взрослого мужчины составляет от 35% до 40% (у атлетов более 50%) веса тела, у женщины несколько меньше (30—35%), у новорожденного еще меньше (20 —- 22 %), у старика — 30 % и меньше (25—27%). Мышечная масса не однородна и распадается на большое число (приблизительно 400) отдельных образований—мускулов, или иначе—мышц. Каждому мускулу свойственны известная величина, форма, отношение к окружающим частям и определенная функция. Рыхлая клетчатка, расположенная внутри мускула, называется endomysium*; она переходит в тонкую пластинку, к-рая одевает мускул снаружи—perimysium** (рис. 2иЗ). Залегающие в endomysium кровеносные сосуды весьма многочисленны и образуют петли, вытянутые по длине мышечных пучков. Мышцы настолько богаты сосудами, что при выключении главных артериальных стволов (напр. при перевязках) окольное кровообращение развивается гл. обр. за счет мышечных артерий. Нервы мускул получает как чувствительные, так и двигательные. Мускул является как бы концевым аппаратом двигательного нерва; вместе взятые они составляют одно целое, одну двигательную единицу. Очень часто нерв и сосуды вступают в мускул в определенном месте—нечто вроде ворот (nilus), area nervo-vasculosa (рис. 4). В огромном большинстве случаев мускулы соединяют части скелета— кости, к-рые при сокращении мускула сбли- * Perimysium internum. ** Perimysium externum. жаются; при этом одна из костей обыкновенно не меняет своего положения; здесь находится укрепленная точка (punctum f ixum) и начало мышцы (origo). Вторая кость совершает движение, почему на ней различаются подвижная точка (punctum mobile) и прикрепление мышцы (insertio). Б. ч. на обо-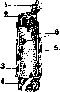
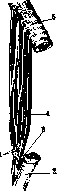
Рисунок 4.
Рисунок 2. Рисунок 2. Мышечное волокно (сильно увеличенное): 1— фибрилы,- 2 — иоле Конгей-ма; з—perimy-sium; 4~sarco-lemma; 5—соединительно- рис з тканное ядро; 6—ядра мышечных волокон. Рисунок 3. Мышца между двумя костями: 1—сухожильные волокна; 2 и 5—периост; з—bursa subtendinea; 4— perimysium externum. Рисунок 4. Area nervo-vasculosa (m. rectus fe- moris): 2—ramus descendens a. circumflexae femoris lateralis; 2 и 3—боковая и медиальная часть; 4—вена; 5—нервная ветвь; в—нерв. их концах мускула имеется сухожилие (ten-do), построенное из плотной соединительной ткани, пучки к-рой переходят в надкостницу. Сухожилия широких мышц имеют форму пластинки и носят название апоневроза (aponeurosis); у длинных мускулов они приближаются к цилиндру. Вспомогательные аппараты мышц представляют различного рода образования соединительнотканного происхождения, так или иначе облегчающие работу мускулов. Сюда относятся фасции, синовиальные влагалища, слизистые и синовиальные сумки, блоки и сесамовидные косточки, фиброзные кольца, retinacula и пр. У лицевых мышц, заложенных б. ч. в жировой клетчатке, фасции и сухожилия отсутствуют. Если мышцы расположены в несколько слоев, то фасция делится на пластинки, или листки, которые друг с другом могут соединяться т. наз. межмышечными перегородками (septa intermuscularia). Фасции дают начало и прикрепление многим мускулам, примыкая в этом отношении к скелету. По Барделебену (Bardeleben), 2/з мышц человеческого тела тесно связаны с фасциями, т. е. их волокна или начинаются или прикрепляются на фасциях. Облехсая мышцы плотным футляром, фасции удерживают их от бесполезных передвижений в сторону. Фасции имеют значительный практический интерес: от фасции в известных случаях зависит тот или иной ход распространения пат. процессов. В определенных местах (при переходе с голени на стопу, с предплечья на кисть) фасции образуют утолщения в виде так называемых ложных связок*; последние, перекидываясь * Ложными связками называются также удвоения серозных оболочек, запустевшие сосуды. между костными выступами, удерживают сухожилия мускулов в их положении. Именно в таких местах развиваются синовиальные каналы сухожилий (vaginae tendinum synoviales). Это—замкнутые с обоих концов тоннели различной длины, в которых в том и другом направлениях движутся сухожилия, притом с минимальным трением, т. к. внутренние стенки каналов и поверхности самих сухожилий выстланы гладкой скользкой оболочкой. В щелевидной полости каналов содержится минимальное количество клейкой жидкости. В одних каналах проходят одиночные сухожилия, в других— по два и более. Такое же значение (уменьшение трения, облегчение работы мышц) имеют слизистые сумки (bursae mucosae) (рис. 3). Последние возникают в утробном периоде, располагаются в пунктах наибольшей подвижности сухожилий обыкновенно вблизи прикрепления мускулов, между сухожилием и костью; находящаяся здесь промежуточная соединительная ткань разрыхляется, и получается щелевидная полость с гладкими стенками. Органы движения представляют сложную систему рычагов. По типу рычагов построены почти все костно-мышечные аппараты; только в одних тип рычага выражен яснее, в других он является более усложненным. Одним из условий действия каждой мышцы является относительная неподвижность одной из точек ее прикрепления. Т. о. каждый предыдущий рычаг должен быть опорой последующему и т. д. В человеческом теле встречаются рычаги как второго рода, так и первого. Рычаги первого рода в организме встречаются довольно часто, их называют рычагами статики, равновесия. Точка опоры в них находится между точками приложения силы мышечной тяги и силы сопротивления. Подобный случай можно наблюдать в сочленениях таза с бедренными костями, между позвонками, в суставе головы с позвоночником (articulatio atlanto-occipitalis); здесь точка опоры (а) лежит на фронтальной оси articulationis atlanto-occipitalis, сила (мышцы затылка, идущие от позвоночника к затылочной кости) прилагается кзади от точки опоры (Pj), а сопротивление (тяжесть го-. ловы) помещено кпереди от нее (Р) (рисунок 5). При многих движениях имеет место рычаг 2-го рода? где точка приложения силы находится между точкой опоры и точкой сопротивления (рис. 6 и 7); так, при сгибании в локтевом суставе точка опоры лежит на фронтальной оси articulatio cubiti (а); неподалеку от нее, в области верхнего отдела предплечья, помещается точка приложения силы (Q) (место прикрепления сгибателя,напр. m. brachialis), а сопротивление (центр тяжести ^предплечья и кисти) располагается значительно дистальнее (Р)*. Рычаг этого типа известен под названием р ы-
Рисунок 5.
* Если кисть при этом поднимает груз (рис. 7), то центр тяжести при значительном отягощении будет располагаться в области груза.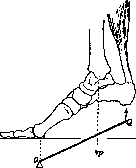
чага скорости, т.к. движения здесь могут совершаться с большой быстротой. На работу мышцы в первую очередь влияют следующие моменты: толщина и длина мышечного брюшка, устройство сустава, расстояние между последним и местом прикрепления мускула и т. д. Напр. сила мускула зависит от его толщины, степень же укорочениямускула (и следовательно высота, на к-рую поднимается груз)—от длины брюшка (см.Мышцы, основные механические явления).
III. Классификация мышц. Все многообразие форм скелетных мышц в теле человека можно объединить в три основные группы—длинные, короткие и широкие мышцы. Кроме того могут быть еще обособлены в отдельную группу т. н. круговые мышцы. Эти последние располагаются вокруг естественных отверстий тела и называются сфинктерами (сжимателя-ми). Длинные мышцы, чаще с веретенообразной формой мышечного брюшка, встречаются обычно на конечностях, т. е. там, где части скелета представляют собой длинные рычаги, производящие больш. дуги движения. Известно, что объем Ркс. 6.' движения зависит помимо отношений между собой ямки и головки в суставе, также и от длины мышцы и костного рычага. В тех же отделах скелета, где рычаги коротки и дуги движения весьма невелики, встречаются короткие мышцы (напр. короткие мышцы спины и затылка). Широкие мышцы представляют собой пластинки. Толщина их незначительна. Встречаются они гл. обр. на туловище (широкая мышца спины, межреберные мышцы, широкая мышца живота). Две последних группы (мышцы груди и живота) своей массой составляют стенки указанных полостей тела. Сухожилия широких мышц чаще имеют характер также широкой пластинки, назыв. сухожильным растяжением, апоневрозом. Кроме приведенной классификации мышц их еще можно подразделить на простые и сложные.— Простые мышцы состоят из одного мышечного брюшка, головки и хвоста. Ход их мускульных пучков довольно однообразен. Наряду с такими мышцами встречаются и более сложные. Усложнение в таких случаях выражается в увеличении числа брюшков (многобрюшные мышцы), числа головок (мышцы двуглавые, трехглавые и более) и наконец числа хвостов. Усложнением формы мышца достигает большого разнообразия в своей деятельности. В дву- или многобрюшных мышцах брюшко (venter) бывает разъединено промежуточным сухожилием (tendo intermedius) или несколькими поперечными сухожильными перемычками, определяющими число брюшков, или прослойками соединительной ткани, имеющими сухожильный характер (inscriptiones tendineae). Эти перемычки срастаются в мышцах с окру- жающим их фиброзным футляром (fascia propria), что создает своего рода опору каждому из этих брюшков в отдельности. Последнее в свою очередь дает возможность изолированного сокращения их и тем самым разнообразия в работе. Примером двубрюшных мышц у человека могут служить mm. biventer, ото -hyoideus, многобрюшных— т. rectus abdominis. Кроме того бывают также мышцы, у к-рых брюшко состоит из нескольких отдельных,соединенных между собой сухожильными прослойками, продольных мышечных конусов, клиновидно внедряющихся друг в друга (напр. дельтовидная мышца—m. deltoideus). Мышцы с несколькими головками имеют несколько обособленных пунктов начала на одной или нескольких костях. К таким мышцам напр. относятся двуглавый мускул плеча, начинающийся от различных участков лопатки (т. biceps brachii), трехглавый мускул плеча (т. triceps brachii), трехглавый мускул голени (m. triceps surae) и др. Далее мышца может начинаться т. наз. зубцами (напр. зубчатые мышцы спины, груди, косые и поперечные мышцы живота). Различное направление хода мускульных пучков в нек-рых из таких зубцов делает их различными между собой и по функции и тем самым вносит разнообразие в работу всей мышцы в цел ом. Мышца, делясь на несколько хвостов, распределяет т.о. силу своей тяги на несколько точек прикрепления, подходя к двум или более костям, причем такая мышца сокращением своего брюшка вызывает или одновременно действие всех своих сухожилий (хвостов) или, напротив, изолирует работу каждого из них в отдельности. Примером таких «ловких» мышц могут служить общие сгибатели пальцев кисти, общие разгибатели кисти. На ноге развитие изолированных сокращений гомологичных мышц выражено слабее. Процесс диференциации такого рода мышц в филогенетическом развитии может дойти до полного выделения части мышцы в самостоятельную единицу. Примером этого может являться мускулатура большого пальца кисти у человека и некоторых обезьян. У животных, ниже стоящих по своей организации и не обладающих способностью тонких движений кистью, мышцы большого пальца не представляют собой отдельных мышечных тел, а являются частью общей мускулатуры всех пальцев. Так, у обезьян т. flexor pollicis longus еще не отделяется от flexor digitorum communis, что впрочем как регрессивная аномалия наблюдается и у человека.—Борелли (ВогеШ; 1710)ввелеще понятие о т. н. перистых мышцах. Чаще в мышцах направление мышечных волокон соответствует направлению волокон в сухожилии. Однако есть и такие мышцы, в к-рых мышечные пучки идут косо и прикрепляются к сухожилию под острым углом. При этом мышечные пучки подходят к сухожилию или с одной стороны (полуперистыемышцы—mm. unipen-
Рисунок 7.
nati) или с двух сторон (перистые или дву-перистые мышцы—mm. Ыреппа1п).Примером первых могут служить m. subclavius и flexor hallucis longus (сухожилие в них располагается сбоку мышечного брюшка), примером вторых—m. rectus femoris и другие мышцы. В последнем случае сухожилие лежит внутри или посередине на поверхности мышцы. Разнообразный характер направления мышечных волокон в мышце и различное отношение их к сухожильной части усложняют определение равнодействующей мышечной тяги и приложения ее к рычагам. По почину Рише (М. Ch. Richet; 1879) физиологи и некоторые анатомы стали еще различать мышцы сильные, преимущественно выполняющие функции статики, равновесия различных частей тела, и мышцы ловкие— динамические. Те и другие отличаются между собой по своемучмакроскопич. построению, способу отношения к рычагам и даже микроскоп. строению (Ed. Retterer, A. Lelievre). Каждая мышца в аппарате костно-мы-шечной системы является той активной силой, к-рая приводит в движение кости, направление же движений зависит от характера соединения данных костей между собой, т. е. от формы суставных поверхностей. Отсюда следует, что положение и группировка мышц (топографическая и фнкц.) должны находиться в определенных соотношениях с устройством суставов, числом осей в них и существующими степенями свободы движений. Зная устройство сустава, можно заранее определить характер и расположение мышц возле него и наоборот—на основании исследования мышечной деятельности—форму того или иного сустава, число осей в нем и пр. Эти взаимоотношения функциональны и весьма динамичны. Всякое нарушение деятельности той или иной группы мышц, особенно в детском возрасте (в период формирования и роста кости), влечет за собой уклонение в нормальном ходе развития формы суставных поверхностей. Из сказанного ясно, что движение животного, следовательно функция мускулатуры, моделирует и детализирует форму суставных поверхностей, возникающую в утробном периоде, и тем определяет степень свободы движения в суставах.—Все мышцы тела по их отношению к рычагам и суставам могут быть подразделены на мышцы, переходящие только через один сустав, и мышцы, переходящие через два или несколько суставов (одно- и многосуставные мышцы). Односуставные мышцы действуют на один сустав, т. е. на одну кость, на один рычаг. Многосуставные мышцы, действуя на несколько рычагов, представляют собой уже этим самым сложную систему. Возьмем для примера нижнюю конечность человека. Мускулатура всей системы ее рычагов распределяется так, что имеются мышцы отдельных суставов и мышцы общие, принадлежащие нескольким суставам. Есть мышцы, к-рые соединяют рычаги только свободного отдела конечности, и есть такие, к-рые соединяют этот свободный отдел с костями пояса и даже туловища. Действие последних мышц сложно. Они приводят в движение не только те кости, к к-рым имеют непосредственное отношение, но из- меняют положение и промежуточных звеньев этой цепи рычагов. Количество мышц в той или иной области может быть настолько велико, что они располагаются в несколько слоев. Так, есть мышцы поверхностные и глубокие. И те и другие могут в свою очередь подразделяться и дальше. Помимо мышц, действующих непосредственно на те или иные суставы между костями скелета, в теле человека и других позвоночных животных встречаются и такие мышцы, к-рые не имеют прямого отношения к суставам. К подобного рода мышцам может быть отнесена мускулатура, располагающаяся выше и ниже подъязычной кости, мышцы дна ротовой полости, промежности, мимическая мускулатура. Что касается взаимоотношений мышц и скелета, то еще Вирхов указывал, что мышечная деятельность является формообразующим фактором в отношении костей, причем не только форма кости, но даже рельеф наружной поверхности кости в значительной мере определяется действием силы мышечного сокращения, resp. мышечной тяги. В местах прикрепления крепких мышечных сухожилий на поверхности костей отмечаются бугры, шероховатости, отростки. С другой стороны, мышцы своим брюшком могут производить боковое давление на поверхность кости, вызывая образование желобков, вдавлений, ямок. Степень развития подобных мышечных отпечатков на костях находится в прямой зависимости от степени развития мышц. Они выражены тем резче, чем сильнее развита мускулатура. Кости людей с сильной мускулатурой отличаются более резко выраженными мышечными отпечатками. Специальный морфол. анализ этих взаимоотношений вскрывает целый ряд весьма интересных деталей. IV. Вариации мышц. Вариации в мышечной системе весьма распространены. Почти на каждом трупе можно найти ту или иную аномалию мышц, а иногда даже сразу несколько. Характер этих вариантов очень разнообразен. При этом могут быть уклонения в форме мышц, в положении начала и прикрепления их, в размере, числе и т. д. Многие исследователи занимались вопросом о происхождении этих аномалий. Нек-рые полагали, что в возникновении их может играть роль раса. Тестю (Tes-tut) и др. отвергают это предположение. Из всех классификаций наиболее заслуживающей внимания до последнего времени считалась классификация Видерсгейма (Wie-dersheim). Все варианты мышц им подразделяются на 3 группы, и возникновение их объясняется с филогенетической точки зрения. К первой группе относятся т. н. регрессивные мышцы—такие, к-рые у человека в норме бывают слабо выражены,- напр. т. pyramidalis, мышцы ушной раковины. Наличие у человека сильно выраженных означенных мышц и будет указывать на регресс в мускулатуре данного отдела. Ко второй группе Видерсгейм относит мышцы атавистического характера, т. е. случаи, когда мышца, встречающаяся как вариант у человека, существует нормально у животного из низших позвоночных (напр. m. sternalis). Встречаясь у человека, они как бы иллюстрируют собой возврат к прошлому. Третья группа—прогрессивные аномалии. Существование этого рода вариаций намечает известные перспективы будущей мускулатуры человека. К таким мышцам можно отнести напр. лишние мышцы пальцев у человека (чаще на руке);сильное развитие мимических мышц также может свидетельствовать о шаге вперёд в их диференциации. По мнению нек-рых авторов вариация есть результат действия какого-либо фактора в стадии эмбриональной жизни. Зная постепенное развитие мышцы, возможно представить себе, по мнению этих авторов, чем обусловлена вариация—простой ли остановкой развития или более глубоким изменением зачатка. Расовые, возрастные, половые, проф. особенности мускулатуры. Вопрос о расовых особенностях мышечной системы представлен в ряде отдельных, не систематизированных наблюдений, описанных в разных анат. и антропологических журналах. Как известно, помимо костного скелета форма туловища находится в заврюимости от развития мышц и подкожножирового слоя. Учение о форме «мягких частей» вообще (подкожножировой слой и мышцы) составляет одну из интересных глав соматологии. В этом отношении в науке имеется богатый материал с точки зрения его расового освещения. Можно указать хотя бы на интересный пример расовых особенностей губ. Дело касается их слизистой части («Schleimhautlippen»), развитие к-рой не одинаково у различных рас. У некоторых это выпячивание слизистой оболочки представляет настоящее выпадение ее. Декворт и Гаушильд (Dackworth, Hau-schild) причину этого усматривают в выпячивании самого нижнего из пучков m. orbi-cularis oris. Этот выворот m. orbic. oris у шимпанзе слабо выражен. Резче он развит у меланезийцев, особенно же у европейцев и негров, причем у последних сильнее. Вообще область изучения внешних форм тела в расовом отношении с точки зрения участия в образовании их именно тех или "других мышц в наст, время является еще мало разработанной.—В озрастная анатомия мышечной системы также представляет собой больше вопросов,-чем фактических данных. Известно, что ребенок рождается уже с вполне диференцированной мышечной системой. В дальнейшем происходит лишь интенсивный рост мышц, к-рый выражается в увеличении длины и толщины первичных мышечных пучков. Для мышечной ткани детского возраста характерно богатство ядрами ее клеточных элементов. Мышцы людей старческого возраста характерны атро-фическими изменениями их мышечных элементов (atrophia senilis). Сила таких мышц истощается, последние становятся дряблыми, вялыми. — Что касается половых различий мышц человека, то по этому вопросу в литературе встречаются лишь краткие замечания самого общего характера. Во всяком случае скелетная мускулатура женщины морфологически не отличается от мышечной системы мужчины. Вследствие особенностей развития подкожножирового слоя у женщины трудно бывает заметить мышечный рельеф ее тела. Известно напр., как хорошо выделяются мышцы на теле мужчины, особенно занимающегося упражнением своей мускулатуры. Мышцы женщины вообще слабее мужских. До сих пор мало заботились о физ., развитии девочек по сравнению с мальчиками. Это могло послужить одной из причин относительной мышечной слабости женщины. Это находит себе отражение также на рельефе женского костяка. Кости женщины менее бугристы, они в меньшей мере, чем мужские несут на себе отпечаток действия силы мышечного натяжения. Однако нередко встречаются женщины с достаточно сильно развитой мускулатурой. В данном случае общие конституциональные свойства, а также проф. и бытовые условия жизни имеют большое значение. Различные виды одежды женщины могут деформировать и уродовать ее мускулатуру. Напр. тугое шнурование талии вызывает деформацию ее тела. Вследствие тугого шнурования мышц живота рельеф последнего нарушается. Живот свешивается вниз и принимает форму отвислого. Этот фактор также деформирую-ще сказывается и на мышцах спины. Проф. особенности мышечной системы человека представляют собой также еще предмет только-что начатых в этом направлении исследований. Пока можно найти лишь самые общие указания на сильно развитые мышцы атлетов, борцов и других лиц, специально, по роду их профессии, упражняющих свою мускулатуру; интенсивная мышечная деятельность, физ. труд ведут к усилению процессов обмена веществ в мышечной ткани. В результате повышенной фнкц. нагрузки структура мышцы функционально изменяется. Объем увеличивается, наступает так наз. рабочая гипертрофия. Не подлежит сомнению и противоположный факт, что напр. у работников умственного труда при отсутствии физ. упражнений тела и вообще ограничений движения мышцы становятся дряблыми, объем их уменьшается. В наст, время в связи с повышением интересов к вопросам физкультуры и рационализации труда изучение проф. особенностей мышечной системы в фнкц. аспекте представляет одну из весьма важных и современных задач науки.—Не все мышцы развиваются равномерно и одинаково интенсивно. Мышечные волокна сильно утолщаются до 3-го месяца утробной жизни; у новорожденного они приблизительно в 5 раз толще, чем у плода 2 мес, а у взрослого во столько же раз толще, чем у новорожденного. В периоде лактации (питания молоком матери) быстро растут брюшные мышцы; в более позднем детском возрасте начинает увеличиваться масса мышц языка, нёба, глотки, жевательных мышц в связи с усиленной функцией соответствующих органов. При рабочей гипертрофии мышц волокна их утолщаются, но количество их может повидимому даже уменьшаться. Регенерация мышц может происходить и в постэмбриональном периоде. V. Методика исследования мышц на трупе. Способ консервирования мышечных препаратов. Для исследова- ния мышц на трупе пользуются методом анат. препаровки. Еще ранее, до рассечения кожи и подлежащих ей слоев подкожной клетчатки и фасций, изучается соответствующая область путем осмотра ее и пальпации через кожу мягких и плотных частей. Весьма полезно при этом научиться проецировать те или иные мышцы на кожу. Разрезом кожи и фасции создается свободный доступ к мышце. При подробном изучении отдельно препарируют фасции. В целях долгого сохранения мышечных препаратов трупы до их препаровки консервируются. Для учебных занятий в секционном зале аиат. институтов не требуется консервирования трупа на долгие сроки. В этих случаях достаточно бывает простых и сравнительно дешевых способов для получения желаемых результатов (см. Бальзамирование тру па). Для консервирования анатом, препаратов вообще и мышечных в частности на долгое время с сохранением их объема и по возможности естественной окраски существовало и ранее очень много различных способов *. Каждый из них имеет свои недостатки и преимущества в том или ином отношении. Способ Шора—см. Бальзамирование трупа. В 1916 г. Лысенков опубликовал свой способ консервирования препаратов без жидкости, с сохранением их объема, путем пропитывания их гигроскопическими веществами. Он нашел, что лучшими для этой цели оказываются глицерин с уксуснокислым калием (Kalium aceticum). Эта смесь напоминает собой жидкость Мельникова-Разведенкова, отличаясь от нее своей концентрацией. Жидкость Лысенкова имеет след. состав: глицерина 500,0, простой воды 1 000,0, уксуснокислого калия 500,0 (по весу), формалина 40,0. В этот раствор кладутся или свежеприготовленные препараты или же предварительно фиксированные формалином на срок до 1 мес, смотря по величине. Для ускорения хода бальзамирования рекомендуется до помещения препарата в раствор инъици-ровать его сосудистую систему указанной жидкостью. Когда препараты достаточно пропитаются жидкостью, их вынимают и хранят в герметически закрытых сосудах или просто под легким прикрытием (напр. под стеклянным колпаком). Длительные наблюдения автора за сохранением качества своих препаратов (нек-рые из них находились под наблюдением до 20 лет к моменту опубликования способа) дают основание считать его метод заслуживающим большого внимания. Мышечные препараты, консервированные по способу Лысенкова, служат образцом анатомической техники в этом отношении. Они весьма удобны в целях преподавания учащимся, а также для хранения как музейных Экспонатов. В. Тонкое, В.. Долго-Сабуров. VІ. Биомеханика М. с. человека. 1. Основные принципы системна го расположения мышц человека. Возникновение и развитие скелетных двигательных аппаратов позвоночного идет всегда по пути совместного развития эле- * Известны методы приготовления т. наз. «сухих препаратов», «мокрых», «влажных», заключенных в жидкость, и т. д. ментов, работающих на сжатие (кости, хрящи) и работающих на растяжение (мышцы, сухожилия, связки). В этом отношении механизм скелетной системы позвоночного принципиально отличается от такового у насекомых или ракообразных. У этих животных панцыри, облекающие отдельные звенья,— вполне жесткие сооружения, работающие как на сжатие, так и на растяжение, так, что на долю мышц выпадает одна только чисто двигательная функция. В противоположность этому у позвоночных мышцы—не только двигательный, но и необходимый структурный элемент, без к-рого прочность всего скелетного сооружения мало отличается от нуля; без отчетливого понимания этой особенности мышечный скелет человека вообще невозможно схематизировать и объяснить. Наоборот, если рассматривать М. с. человека не изолированно, а в тесной связи со всей совокупностью конструктивных элементов скелета и притом уделять решающее-внимание именно статической, а не двигательной роли мышц скелета, то их расположение и строение окажутся очень ясными и хорошо объяснимыми. Основная статическая схема, имеющая место во всех отделах скелета позвоночного, содержит сжатый элемент (кость) в. середине, а растянутые элементы (мышцы и связки)—со всех сторон кругом него. Такое расположение аналогично'расположению мачты с вантами (рис. 8). Статические роли мышц и связок очень близки между собой*, и могут быть поняты при совокупном рассматривании тех и других. Фнкц. различие тех и других начинается там, где появляется возможность движений, осуществляемых.: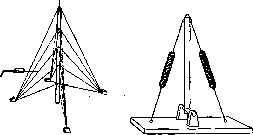
Рисунок 8. Основная струк- Рисунок 9. Схема звена на турная схема скелета по- одноосном сочленении с звоночных—мачта с ван- двумя мышечными рястами (растяжками). Ср. тяжками (стойкими анта-с рис. 16. гонистами).
специально мышцей, а не связкой. Поэтому в истории развития совершенно параллельно-идут процессы специализации сочленений, ограничения их подвижности, и процессы специализации мышечно-связочного аппарата: мышечная ткань заменяется более простой сухожильной во всех тех растяжках, к-рые в силу ограничения подвижности сочленения уже не могут нести двигательных функций и вынуждаются к одной лишь статической роли. Нечто подобное наблюдается и у индивидуумов в случае пат. нарушений подвижности сустава: мышца, утратившая-возможность выполнения движений, не атрофируется совершенно, но сохраняет полностью свои статические функции, превра—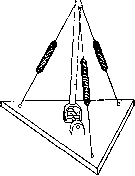
Рисунок 10. Схема звена на двуосном сочленении с тремя мышечными растяжками .
щаясь в сухожильноподобную растяжку.— Итак среди растяжек, окружающих сжатый элемент и его сочленение обязательно со всех сторон, мышечные свойства сохраняются только у некоторых в зависимости от условий и возможностей подвижности сочленения. Встречающиеся здесь случаи таковы. В одноосном сочленении подвижность возможна только в одном из двух направлений (рис. 9); в таком сочленении мы должны ожидать двух мышц, роли которых по необходимости прямо противоположны. Это т. н. мышцы - антагонисты (см. Движение); точнее называть их стойкими или постоянными антагонистами. Все остальные растяжки при таком сочленении превращены в связочный аппарат. Весьма характерным примером такого случая может служить плече-ло-ктевое сочленение (articulatio brachio-ulnaris): фронтально и дорсально оно снабжено двумя мышцами-антагонистами (m. brachialis int. и медиальная и лятеральная головки m. tricipitis br.) и двумя связками (septa intermuscularia brachii и lig. lateralia). Очень распространено ошибочное представление о том, что по тем же соображениям двухосное сочленение требует по меньшей мере двух пар антагонистов, трехосное сочленение—трех пар. Это мнение не только основано на недоразумении, но и прямо опровергается действительностью (см. ниже мышцы предплечья). Из рисунка 10, схематически изображающего двухосное сочленение, ясно, что для исчерпывающего использования подвижности такого сочленения ему необходимы всего три мышечные растяжки, причем в каждом из возможных движений одна из них является антагони-. стом к обеим остальным. Точно так же трехосному сочленению необходимы и достаточны всего на всего четыре мышечные растяжки (рис. 11). Действительно этот рисунок показывает, что различные комбинации этих четырех мышц обеспечивают возможность вращения костного звена около трех взаимно перпендикулярных осей АА, ВВ и СС: мышцы 1 и 2 против 3 и 4 дают вращение вокруг оси АА, мышцы 1 и 4 против 2 и 3—вокруг оси ВВ, мышцы 1 и 3 против 2 и 4—-вокруг оси СС. В обоих последних случаях (рис. 10 и 11) мы уже не имеем дела со стойкими антагонистами; здесь, смотря по роду движения, каждая пара мышц может оказаться фнкц. антагонистами или агонистами друг для друга. Из сказанного следует еще и то, что сочленения с меньшим числом осей подвижности имеют более мощный связочный аппарат, нежели трехосные сочленения (со- членовная сумка лопаточно - плечевого сочленения не содержит ни одного вспомогательного сухожильного пучка, и все сочленение держится на мышцах), а также то, что связки расположены как-раз на тех сторонах сочленений, к-рые свободны от мышц.— Изложенные сейчас соображения о минимально необходимом количестве мышц не оправдываются для весьма важных механических узлов скелета—лопаточно-плечевых и тазобедренных сочленений, имеющих более чем по 4 мышцы. Это объяснимо и биологически и механически. Биологически (см. ниже) эти пункты являются местами встречи самостоятельных мышц конечностей и туловищных мышц поясов, развивающихся из разных источников навстречу друг другу. Механически это объясняется тем, что мышцам плеча и бедра приходится не только двигать и поддерживать самые эти звенья (это их количественно наименьшая нагрузка), но и служить опорой для далеко отстоящих дистальных звеньев этих конечностей. Кисть руки не работает, когда нагружено плечо, но плеяо работает, когда нагружена кисть, и притом нагрузка плечевых мышц в этом случае (за счет большей длины рычагов) в'5—6 раз больше нагрузки мышц самой кисти. В связи с этим в узловых сочленениях между конечностями и туловищем помещаются не только мышцы, обеспечивающие движения в самих этих сочленениях, но еще и весьма мощные мышцы, обеспечивающие возможность движений и фиксаций на периферии. Интересно, что в плечевом сочленении мышц того и другого назначения по 4 штуки (см. ниже). 2. Библ. схема мышц туловищной трубки и поясов. Туловищная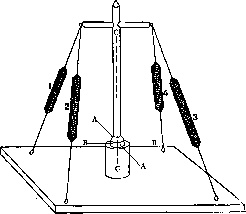
Рисунок 11. Схема звена на трехосном (шароЕом) сочленении с четырьмя мышечными растяжками.
костно-мышечная система человеческого тела сохраняет на всю жизнь сегментарное строение. Мышечные сегменты, имевшие первоначально вид, подобный гребенке, сохраняют его и у взрослого человека в области грудной клетки (mm. intercostales), заменив однако первоначальные сухожильные межсегментные перемычки костными (ребрами). То же .сегментарное строение выявляется иногда довольно ясно и в области брюшных и спинных мышц (напр. m. rhomboideus scapulae). Мышцы каждого сегмента эмбриологически очень рано разделяются на спинную и брюшную группы; последняя в свою очередь расслаивается на три слоя (рис. 12). Главное осложнение сегментарной - rev™, схемы представляют с$$^ \^*~~***^Ь11Щ сооои верхний и ниж->--=*'==-* ний пояса конечно- стей, построенные по вполне сходному плану. Однако вполне одинаковое поперечное расположение передних и задних конечностей (локтями и коленями наружу), свойственное пресмыкающимся, заменяется у млекопитающих поворотом обе-Рисунок 12. схематический их пар конечностей разрез мышц туловищ- в сагитальную ПЛО- НОЙгоР шКоИBnr°a3uBsT)4H°" скость. При этом колени поворачиваются вперед, а локти назад (1-й поворот); кроме того, чтобы кисти передних конечностей не оказались повернутыми кзади, происходит еще перекручивание обеих костей предплечья в положение постоянной пронации {2-й поворот). Механизм супинации появляется только у антропоидов (см. Обезьяны). Схема расположения конечностей вполне уясняется рисунком Брауса (рис. 13). Мышцы поясов возникают двояким образом. Одни из них начинаются на проксимальной кости конечности (плече, бедре) и развиваются по направлению к костям лояса; это — самостоятельные мышцы конечностей (autoch-tone Muskelblastem, Braus); они, как и сами конечности, эмбриологически принадлежат к вентральной стенке тела. Другая группа мышц начинается от туловища и растет навстречу первым, срастаясь С КОСТЯМИ ПО- рис 13 Схема развития ЯСОВ и укрепляя ИХ расположения конечно-K ТУЛОВИЩу; ЭТО — стей; а—расположение у туловищные мышцы пресмыкающихся; Ъ - у J ,-гг- млекопитающих. Прямая поясов (Korpermus- стрелка-продольная ось CUlatur, Braus), ВОЗ- тела, ее острее — голов- никающие ИЗ дор- ной конец- Маленькие ^Г rt >■ стрелки—основные оси Сальной Стенки тела суставов, у передних ко- зародыша. ПроИСХО- нечностей показан схе- ДЯ ИЛИ ТОЛЬКО ИЗ матически ход п. radia- о Us, у задних—п. lschia- вентральнои или то- dlci *и п^ per0naei. (По ЛЬКО ИЗ дорсальной Braus'y.) стенки, каждая из этих двух групп образует однако по два пучка, функционально являющихся вентральным и дорсальным. Самостоятельные мышцы верхней конечности образуют прежде всего пару коротких пучков с брюшной и спинной •стороны плеча (рис. 14,1 и 2); у человека это— mm. infraspinatus и supraspinatus. Далее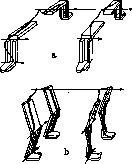
возникает вторая, более длинная пара самостоятельных мышц (рис. 14, 3 и 4); эта пара частью заходит за пределы костей пояса и укрепляется к костям туловища—ребрам и позвоночнику. У человека это—-m. pecto-ralis major и т. latissimus dorsi. Группа туловищных мышц поясов вдвигается между сегментными мышцами туловища (рис. 14, 5 и б) и самостоятельными мышцами верхней конечности. У человека две мышцы этой группы (m. rhomboideus и т. serratus anterior) поддерживают лопатку (рисунок 14, 7 и 8) и одна (m. subclavius) (рис. 14,"9)— ключицу.
3. М. с. позвоночного столба н туловища. Выше было указано, что на туловище человека кроме собственно туловищных мышц расположены еще многочисленные мышцы поясов конечностей. Мио-статический смысл самостоятельных мышц Рисунок 14. Схематический разрез туловищной трубки позвоночного с изображением хода развития мышц поясов. Из мышц показаны собственные мышцы туловища: 5—m. inter-cost.int., 6—т. intercost. ext.; самостоятельною мышцы конечностей (развиваются от конечностей к туловищу): 1—т. brachialis superior; 2—т. bra-chialis inferior; 3— т. pectoralis major; 4— т. latissimus dorsi; туловищные мышцы поясов (развиваются от туловища к конечностям): 7—m. rhomboideus; S—т. serratus ant.; 9—т. subclavius. (По Braus'y.) туловища уясняется лучше всего из рассмотрения их у четвероногих млекопитающих. Позвоночный столб имеет у последних строение арки (рис. 15), т. е. сооружения, у которого сжатая,часть расположена наверху и имеет выпуклость, обращенную кверху; растянутая часть сооружения (затяжка арки) натянута между пятами арки, препятствуя их раздвиганию в стороны. Арка позвоночника четвероногого млекопитающего укреплена двояким образом. Во первых она снабжена затяжкой, к-рой первоначально служила длинная парная мышечная лента, тянувшаяся от лобкового сращения по вентральной стороне туловища до самой нижней челюсти (рис. 15, А). Впоследствии эта лента была вытеснена в грудном отрезке развившейся здесь грудинной костью, и от нее уцелели только брюшной (mm. rec-ti abdominis) и шейный кусок (mm. sterno-hyoideus, sterno-thyreo-hyoideus, diaphrag-ma oris).—Если бы цепочкообразная дуга позвоночника служила только аркой, то звенья ее—позвонки—должны были бы быть прочно заклинены, как сжатые камни арочного моста. Этим была бы достигнута прочность, однако последствием ее явилась бы жесткость. Между тем позвоночник млекопитающего есть невоспроизведенная еще техникой конструкция гибкой арки. Поэтому сводовая часть этого сооружения построена не по типу жесткой арки, а по упомянутому уже основному типу цепочки с Байтовыми растяжками. Такие растяжки. облекают позвоночную цепочку с четырех сторон: брюш-.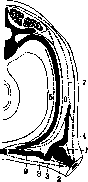
в. м. э. т. xix.
ной, спинной и обоих боков, в связи с чем и каждый из позвонков имеет приближенно крестообразную форму. Брюшная растяжка (lig. longitudinale ant., рис. 15,В)—чисто сухожильная, прочие три пучка—преимущественно мышечные. Остистые и поперечные
Рисунок 15. Схема продольных растяжек туловища четвероногого (собаки): А—брюшная затяжка; В—lig. longitud. anterius; С—tractus medialis m. erectoris trunci.
отростки позвонков в этой схеме следует рассматривать как окостеневшие прослойки между мышечными сегментами; однако у человека в позвоночных растяжках (mm. longissimi dorsi) большая часть волокон претерпела слияние сегментов и тянется не через 1, а через 2 — 6 сегментов. — Весь массив задних и боковых растяжек (mm. erec-tores trunci, s. longissimi dorsi), раздвоенный на два кабеля, тянется вдоль всей спины человека—от крестца и подвздошных костей до затылочной кости. Задняя, или спинная растяжка (рис. 15, С) включает в себя mm. interspinales и spinales (рис. 16, А и В), идущие вертикально, и mm. rotatores, mul-tifidi, semispinales (рис. 16, С), идущие наклонно, в полном подобии с рис. 8. Боковые растяжки соединяют между собой поперечные отростки позвонков (mm. intertransver-sarii, отчасти m. quadratus mmborum, рис. 16, D), отчасти же связывают другую разновидность межсегментных прослоек—ребра (mm. ilio-costales, рис. 16, JE). В грудной части позвоночника, обращенной выпуклостью к спине, как настоящая арка, прочность достигается вполне уже брюшной затяжкой (рис. 15, А), и спинное оборудование используется только для гибкости; в связи с этим оно чисто мышечное. В поясничном же и шейном отделах, где позвоночник обращен выпуклостью к брюшной стороне, необходимы постоянные статические скрепы: в этих отделах продольные мышцы спины усилены толстыми сухожильными пластинами: fascia lumbo-dorsalis и lig. nuchae; наличие последней объясняется еще тем, что статически шейный отдел представляет собой уже не арку, а консоль с подвесом наверху. Такая статически уравновешенная система несет на себе все туловище, построенное наподобие футляра с гибкими трехслойными стенками;. Эти слои и свойственные им направления волокон прослеживаются на всем протяжении туловища, от подвздошных костей до шеи; в грудном отделе они имеют сегментарное строение, к-рое уже утратилось в брюшном и шейном отделах. Самый глубокий слой имеет трансверсаль-ные волокна и включает в себя mm. trans-versi thoracis и transversi abdominis. Следующий слой имеет косо идущие волокна; он состоит из mm. intercostales interni и mm. obliqui abdominis interni. Наружный слой имеет обратное косое направление волокон. Этот слой самый обширный из всех; он лежит поверхностнее структурных элементов рисунка 15 и перекрывает сзади всю шею> (mm. splenii), большую часть спины (тт.. serrati post., levatores costarum), бока и брюшную сторону туловища (mm. intercost. ext.,. mm. obliqui abdominis ext.), а также боковые стороны шеи (mm. scaleni). Направления слоев широких мышц туловища лучше-всего прослеживаются на рис. 17.—Статическая роль этих слоев определяется вполне направлениями их волокон; динамические функции их чрезвычайно разнообразны, так как ими исчерпываются все мышцы брюшной стенки. Наружный слой, поднимающий и раздвигающий ребра и натягивающий стенки живота, весь может быть использован для вдоха; второй и отчасти внутренний слой пригоден для выдоха (опускание ребер и втягивание брюшной стенки). 4. М. с. плечевого пояса. Структурные схемы плечевого и тазового поясов имеют между собой большое эмбриологическое сходство, но механические условия их функциони-А рования уже у четвероногих млекопитающих очень различны. Прежде всего плечевой пояс представляет собой две в очень гибкие полуцепи, тогда как та- ■ зовой есть одна почти жесткая замкнутая цепь. Как один, так и другой пояс четвероногого можно рассматривать как козлы, в рогатки к-рых положена горизонтальная балка позвоночного столба. В плечевом поясе такую рогатку образует цепь из обеих ключиц, manubr. sterni И связанных С НИМИ ных элементов системы мышц; на этой цепи головной конец туловища лежит так,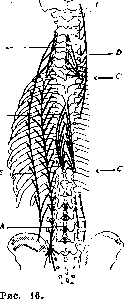
Схема состав-
ш. erector trunci (no Braus'y)- Нанесены только немногие типовые элементы; так напр. уз- как Обучающийсяпла- лы, отмеченные стрел- тчятк пржит тта ттпмп- ками С> в действитель-вать лежит напомо- ностя находятся у каж- Чах, за которые его дого поперечного отрост-подвешивают. Схема ка. сравнить строение передних конечное- этих ^злов с ркс- 8-тей и плечевого пояса имеет таким образом вид буквы М, причем самый пояс аналогичен висячему мосту на двух опорах. В тазовом поясе вместо висячей петли имеется свод, которому хвостовая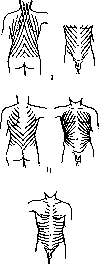
часть позвоночника—крестец—служит замком. Как и подобает замку свода, крестец заклинен между двумя расходящимися кверху сочленовными площадками тазовых костей, образующих т. о. опять-таки рогатку, на этот раз уже более жесткую. Вытекающая из этого различия ,^Jjt[L разница в распределе--J&Wa \ нии усилий в обоих поясах совершенно ясна. Растяжение висячей петли переднего пояса стремится сблизить точки подвеса; такое сооружение для устойчивости нуждается в верхней распорке, работающей на сжатие. Такой распоркой плечевому поясу служит упругий бочонок грудной клетки, на спинную сторону которого давят пластины лопаток, распределяя свое давление на довольно большую поверхность*. Давление на свод или арку тазового пояса стремится раздвинуть его точки опоры—■ тазобедренные сочленения; поэтому этот свод, как и арка позвоночника (см. выше), нуждается в нижней затяжке. Такой затяжкой служит у четвероногих symphysispubis, располагающийся как-раз под позвоночником; у человека в результате прямохождения механические условия работы таза резко изменились (см. ниже).
Как и подобает висячему сооружению, работающему на растяжение, плечевой пояс укреплен почти полностью на мышцах; костный аппарат его очень беден, и как-раз у четвероногих, у к-рых плечевой пояс работает в точности в вышеописанных условиях, ключицы или вполне отсутствуют (копытные) или представлены небольшими хрящевыми вкраплениями (хищные). Стоит отметить, что с переходом на вертикальную стойку плечевой пояс выиграл в смысле подвижности по крайней мере так же много, как тазовой пояс проиграл. Петля плечевого пояса, на которой при четвероногой стойке был подвешен позвоночник, у человека освободилась от нагрузки и смогла полностью использовать всю гибкость, присущую ей как подвесному сооружению. У четвероногого передняя конечность поддерживает туловище, у человека туловище поддерживает переднюю конечность; это разгружает плечевой пояс примерно во столько раз, во сколько рука легче четверти туловища с годовой, т. е. в 3—4 раза. Подвес плечевого пояса четвероногих (рис. 18) есть широкий Рисунок 17. Схема направлений хода волокон широких мышц туловища: I—поверхностный слой; II— средний слой; III— глубокий слой. мышечный бандаж, на к-ром висит грудная клетка. Этот бандаж охватывает грудную клетку с брюшной стороны и с боков и «застегивается» на спинной стороне—у остистых отростков позвоночника. К устоям висячего моста — конечностям — этот бандаж подвешен за спинные (верхние) края лопаток, с к-рыми он сращен по всей их длине; на брюшной стороне он начинается от sternum и от боковых поверхностей ребер. Такой же подвесной бандаж в общих чертах сохранен и у человека, но в связи с уменьшением нагрузки он сильно редуцирован на брюшной стороне. Он состоит дорсально из mm. rhomboidei scapulae, вентрально — из mm. serrati anteriores. Динамика плечевого пояса. Лопатка, пока она прижата к задней стороне грудной клетки, имеет 3 степени подвижности (см. Движения) — приведение — отведение, поднимание —опускание, вращение внутрь и наружу. Сообразно с этим мы ожидаем при лопатке 4 мышечные растяжки (см. выше). Она и имеет их как-раз четыре, расположенных крестом. Две из них, т. rhomboideus и т. serratus ant., ведающие приведением и отведением лопатки, были выше описаны в качестве бандажа; две другие имеют анатомически одно общее наименование—m. trapezius. В связи с переходом на вертикальную стойку у человека роль подвеса перешла к этой второй паре,подвешивающей плечевой пояс к туловищу; первая пара служит у человека преимуществ, для прижатия лопатки к стенке грудной клетки, что у четвероногих достигалось уже весом туловища; фнкц. недостаточность этой пары ведет у человека к образованию scapulae alatae. Все движения лопатки осуществляются за счет перераспределения напряжений в описанных че- * Возможно, что сжатая с боков форма грудной клетки четвероногих обусловлена именно этим боковым давлением лопаток. У человека, не опирающегося на руки, это давление отсутствует, и грудная клетка становится шире в трансверсальном направлении.

Рисунок 19.
Рисунок 18. Схема мышечного бандажа плечевого пояса четвероногого млекопитающего. Рисунок 19. Полусхема расположения и хода волокон подвеса лопатки: 1,2th.3— верхняя,средняя и нижняя порции m. trapezii; 4—т. rhomboideus; 5—т. levator anguli scapulae. тырех подвесках. Приведение выполняется m. rhomboideus и средними порциями т. trapezii, отведение—m. serratus ant. Поднимание лопатки совершает верхняя половина m. trapezii, в соответствии с большей трудностью поднимания более массивная, нежели нижняя половина; кроме того ей помогает еще переселившийся на шею'ку-сок бандажа — m. levator anguli scapulae , (рисунок 19). Опускание совершается в •13 891 МЫШЕЧНА силу тяжести и нижнею половиной m. tra-pezii.—Биомеханически вращение лопатки играет несравненно большую роль во всех движениях плеча, нежели ее поднимание и опускание. Поэтому преобладающая функция верхней порции m. trapezii не поднимание, а статическое поддерживание лопатки. Вращения же лопатки выполняются координированными действиями всех четырех растяжек. Вращение внутрь, соответствующее подниманию руки и потому более тяжелое, осуществляется двумя парами тяг: 1) порциями 1 и 3 m. trapezii (см. рисунок 19) и 2) нижним отделом m. serrati ant. совместно с верхним отделом m. rhomboidei. Вращение наружу, подкрепляемое тяжестью, обеспечено одной парой тяг—верхним отделом m. serrati ant. и нижним отделом т. rhomboidei. Все описанные мышцы возникают на туловище й развиваются от него по направле-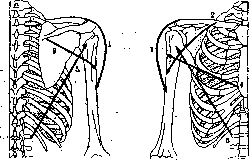
Рнс. 20. Направления тяг длинной группы мышц плечевого сочленения. Слева—вид сзади, справа—вид спереди.
нию к конечностям. Навстречу им от плечевой кости развиваются самостоятельные мышцы верхней конечности, заведующие уже движениями в плечевом сочленении. Разумеется, если положение конечности фиксировано (например хваткой за неподвижный предмет), то те же мышцы способны двигать и лопатку. Рассматривая биомеханически эту труппу мышц, мы еще раз встречаемся с фактом (отмеченным уже при характеристике m. trapezii), что анат. и биомеханические; мышечные единицы могут очень значительно разниться друг от друга. Иногда то, что анатомически есть группа мышц, биомеханически есть одна мышца (напр. т. longissi'mi dorsi); иногда, наоборот, единая анатомически мышца для биомеханики есть целая группа мышц (напр. ,m. trapezius или т. deltoideus). Выше было упомянуто, что самостоятельные мышцы плеча делятся на две группы—коротких мышц, оканчивающихся на костях плечевого пояса, и длинных, более массивных, заходящих далеко на туловище. У человека короткие мышцы составляют законченную группу, длинные же мышцы должны быть для такой же биомеханической законченности пополнены одной из коротких мышц, самой массивной из них (m. deltoideus), функционально не относящейся к группе коротких.—Группа длинных мышц анатомически состоит из четырех мышц,: а'биомеханически—из пяти тяг; схема ее дана на рис. 20. Я СИСТЕМА 392 Разгибание плеча (т. е. движение его вперед—наружу) совершается толстой перистой срединной порцией m. deltoidei, мышцы, которая биомеханически распадается на три самостоятельные единицы. Механически этот разгибающийся пучок заканчивается не на ключице (как анат. m. deltoideus), а продолжается и вверх от ключицы в виде передне-верхнего края m. trapezii. У собак то и другое есть и анатомически единая мышца—levator humeri proprius. Сгибание плеча идет в направлении пучка 5 (рис. 20) m. Iatissimi dorsi, и биомеханически—единой с ней древнейшей части этой мышцы — m. teretis majoris. Направление действия m. Iatissimi привело к колоритному старинному названию ее—m. aniscalp-tor (подтирательная мышца), что не совсем верно, т. к. она ротирует плечо в направлении сверхпронации, т. е. ладонью от спины.—■ Другая пара движений плеча есть анте-версия (поднимание вперед—внутрь) и ретроверсия (опускание назад—наружу). Антеверсия совершается совместным действием пучков 2 и 4 (рис. 20) (передняя порция m. deltoideus и т. pectoralis major). Ретроверсия происходит при помощи пучка 3 (рис. 20), анатомически соответствующего задней трети m. deltoidei и имеющего механическое продолжение над spina scapulae в виде порции 2 (рис. 19) т. trapezii.—Третий вид движений плеча, разгибание и сгибание в фронтальной плоскости, представляет собой смесь обоих первых видов; напр. разгибание этого рода—сочетание разгибания (вперед — наружу) и ретровер-сии (назад — наружу), сообразно с чем используются для этого движения и мышечные тяги описываемой группы.—Наконец р о-т а ц и я плеча вокруг его собственной продольной оси—ротация н а р у ж у или сверх-супинация и ротация внутрь, или сверхпронация, осуществляются пучками 4 (т. pectoralis maj.) и 5 (т. Iatissimi), к-рые для этого движения являются антагонистами. При ротации наружу или сверхсупинации сухожилие m. pectoralis навивается на плечевую кость, как лента на вал, при сверхпронации—разматывается обратно, Необходимо еще вкратце рассмотреть роль элементов плечевого пояса в движениях плеча. Если бы лопатка не была при каждом из этих движений прочно фиксирована, то все описанные только-что мышечн. тяги совершали бы как-раз обратное тому, что было про них сказано: они двигали бы лопатку при неподвижно висящей руке, так как и масса и сопротивление (статический момент) у лопатки значительно меньше, нежели у плеча. Движения плеча и всей руки возможны только при условии содружественного (сиыергетического) напряжения соответствующих мышц лопатки. Биомеханически все мышечные тяги, двигающие плечо и всю руку, должны быть прослежены до действительно фиксированных точек опоры—позвоночника и грудной клетки. Лопатка, к-рая у четвероногих была действительно фиксированным структурным элементом, удерживаемым тяжестью всего туловища, висящего на ней, превратилась у человека фактически в костное вкрапление в толщу ам мышц, связывающих плечо с позвоночнико-вой системой, вкрапление, напоминающее собой функционально примерно коленную чашку. Ясно, что движения плеча не предполагают полной, спастической фиксации всех мышц лопатки. Наоборот, на фоне общего (тонического) фиксирующего напряжения лопаточной мускулатуры с особой отчетливостью (тетанически) напрягаются те мышцы, к-рые двигают лопатку в направлении, содружественном с данным направлением движения плеча. Эти содружественные направления явствуют из след. таблицы. Движения плеча Движения л о-п а т к и Разгибание (вперед—наружу) Сгибание (назад—медиально) Антеверсия (вперед—медиально) Ретроверсия (назад-наружу) Приведение (вращение вперед относительно вертикальной оси) Отведение (вращение назад относительно вертикальной оси) Вращение внутрь Вращение наружу Не участвует Отставание нижнего угла от грудной клетки Отведение Приведение Сопоставляя данные этой таблицы с приведенными выше указаниями на функции мышц лопатки и плеча, легко проследить встречающиеся здесь синергии. Вторая, короткая группа самостоятельных мышц плечевого сочленения вся кроме одной мышцы связана с лопаткой. Все мышцы этой группы лежат сравнительно глубоко и почти целиком прикрыты представителями длинной группы. Четыре мышцы из этой группы, связывающие плечо с лопаткой, образуют и четыре биомеханических тяжа; пятая (т. pectoralis minor) не имеет более связи с плечом (рис. 21). Функции этих мышц характеризуются след. таблицей (цифры соответствуют обозначениям на рис. 21). Наименование Функция Синергет из представителей длинной группы 1. М. supraspinatus 2. М. infraspinatus 3. М. subscapularis 4. М. coraco-brachia-lis Разгибание плеча Сгибание(и сверхсупинация) Сгибание(и сверхпронация) Антеверсия Средняя порция m. del-toidei М. latissimus dorsi (сгибание и сверхпронация) М. latissimus dorsi Передняя порция т. . deltoidei При движениях в плечевом сочленении работают как длинная, так и короткая группы мышц. Очень вероятно (хотя строго еще не доказано), что для несильных, быстрых и точных движений употребляется преимущественно короткая группа; для сильных движений и для фиксаций—длинная группа. Следует особенно подчеркнуть, что одна и та же мышца, состоящая на службе при многоосном сочленении, может оказаться исполни^ тельницей прямо противоположных функций в зависимости от исходного положения конечности. Так напр. m. latissimus dorsi совершает движение сверхпронации, если перед этим плечо было сверхсупинировано, и эта же мышца вращает его в обратную сторону, если исходное положение плеча было полной сверхпронацией. Средняя порция т. deltoidei может способствовать антеверсии плеча, если таковая уже начата другой мышцей, и способствовать ретроверсии,если началась ретроверсия. В силу этого полное описание двигательных функций мышц плечевого сочленения невозможно; в значительной степени разбору встречающихся здесь случаев могут способствовать схематические рисунки 20 и 21. 5. М. система верхней конечности. Мускулатура плечевой кости, так же, как и бедренной, иллюстрирует еще одну биологически важную функцию М. с. Обе мышцы плече-локтевого сочленения— m. brachialis interims и две короткие головки m. tricipitis br. обрастают плечевую кость по всей поверхности ее дистальной половины. Между тем для выполнения движений в плече-локтевом сочленении было бы достаточно точечного прикрепления к костям (наподобие прикрепления m. bici-
Рисунок 21. Направления тяг короткой группы мышц плечевого сочленения. Слева—вид спереди, справа—вид сзади.
pitis br. к tuberculum radii). Такое обрастание— пример укрепления самой кости растяжками на протяжении ее диафиза. Это укрепление имеет реальное значение не только у ребенка при еще не окостеневшей плечевой «кости», но и у взрослого индивида; достаточно отметить, насколько более часты переломы в верхней половине плечевой кости (collum chirurgicum), не укрепленной подобным же образом. Функции обеих упомянутых мышц, ясны из простого анат. описания; однако следует отметить, что и сгибатель и разгибатель локтя имеют явные биомеханические продолжения в области плечевого сочленения. М. brachialis int. начинается у наружного края плеча как-раз там, где прирастает m. deltoideus; вместе с этой мышцей он образует как бы единую мышечную тягу, пришитую посередине к плечевой кости. Подобным же образом медиальная головка m. tricipitis сливается с т. со-raco-brachialiSj механически продолжающим его кверху.—Двуглавая мышца плеча перекидывается через целых три сочленения и нигде не связана с плечевой костью. При фиксации двух сочленений из перекрываемых ею трех мышца эта работает как простая од-носуставная и функции ее несложны. В плечевом сочленении длинная головка m. bicipi-tis вызывает разгибание плеча, короткая— антеверсию,обе головки вместе—поднимание плеча сагитально вперед. На плече-локтевое сочленение в силу особо выгодного расположения мышца эта действует в 3—4 раза сильнее, чем на плечевое (Браус), вызывая сгибание локтя; максимальную силу в этом сочленении она развивает при прямом угле между предплечьем и плечом. По отношению к лок-те-лучевому сочленению эта мышца работает как супинатор и притом значительно более сильный, чем собственные супинаторы предплечья. Интересно указать, что супинация правой руки соответствует по направлению ввинчиванию винта с нормальной (правой) нарезкой; выбор именно правой нарезки в качестве нормальной стоит в несомненной связи с большой супинаторной силой m. bicipitis при полусогнутом локте. Стойким антагонистом этому движению является m. pronator quadratus. Общая сила сгибателей локтя у человека раза в полтора больше, чем разгибателей. Здесь налицо то же явление, что в мышцах лопатки (см. выше): разгибанию способствует, а сгибанию противодействует тяжесть. Строение М. с. предплечья выясняется с большой отчетливостью из анализа подвижности кисти. Система запястных сочленений в общей сложности двухосна, т. е. дает две степени свободы подвижности (см. Движения); локте-лучевое сочленение, обусловливающее про- и супинацию, одноосно, т. е. об-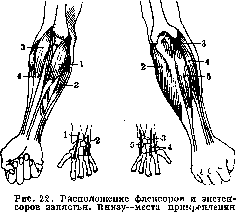
сухошшшй. (По МоШег'у.)
ладает одной степенью свободы. Итого мышцам предплечья, поскольку они смещают луч и кисть, приходится осуществлять три степени свободы движений; в связи с этим они распределяются на4 отчетливо выраженные тяги (рис. 22), разбивающиеся на две парные группы. На вентральной стороне предплечья и у медиального мыщелка плеча помещается группа сгибателей-пронаторов; на дорсальной стороне и у лятерального мыщелка —■ группа разгибателей-супинаторов. Сухожилия обеих этих групп раздваиваются к дистальному концу предплечья и оканчиваются соответственно у лучевого и локтевого краев запястья: сгибательная группа—на' волярной, разгибательная—на дорсальной стороне. Такое расположение весьма близко воспроизводит приведенный выше рисунок 11. Сопоставляя его с рис. 22, легко проверить следующую схему действия этой системы мышц: Наименование мышц : Сила про- и супинационного действия этих мышц невелика и используется лишь как синергетическая помощь про- и супинаторам-специалистам.—-Для характеристики статики кисти необходимо указать еще следующее. Выше указывалось, что в направлении движений, невозможных для данного сочленения, мышечная ткань заменяется связочным аппаратом. Для двухосного лучеза-пястного сочленения невозможным является вращение кисти вокруг продольной оси (заменяющееся про- и супинацией). В прямой связи с этим запястье охвачено мощным сухожильным браслетом (apparatus ligamento sus carpi, рис. 23), преобладающая часть волокон которого идет как-раз в направлении недостающей подвижности кольцеобразно вокруг запястья. Собственные мышцы кисти не требуют особого системного анализа; необходимо внимательнее остановиться только на функциях длинных пальцевых мышц и на их синер-гиях с короткими (mm. lumbricales). Mm. interossei имеют свою специальную простую функцию отведения и приведения пальцев и почти не участвуют в сплошной сгибатель-но - разгибательной синергии. Для анализа движений пальцев следует разделить их все на три типа. Одни из движений возможны свободно-активно, т. е. за счет самостоятельного действия мышц. Другие осуществимы только вынужденно-активно, т. е. при условии использования внешних сил, фиксирующих одни сочленения и не мешающих свободе других. Наконец, нек-рые движения возможны только пассивно—за счет одних лишь внешних сил. Возможные здесь комбинации яснее всего иллюстрируются схемами рис. 24.—Прежде всего необходимо указать, что длинные мышцы пальцев, перекидываясь и через запястные сочленения, вызывают кроме движений пальцев еще и сгибания и разгибания запястья. Как и другая подобная многосуставная группа мыщц (см. ниже), эти мышцы слишком коротки, или, лучше сказать, имеют слишком малую изменчивость длины (сократимость и растяжимость), чтобы одновременно пол- Движение Сокра- Растяги- кисти щаются ваются Сгибание №№ 1 И 2 (3+4) и 5 Разгибание (3 + 4) И 5 1 и 2 Приведение 2 И 5 1 И (3 + 4) Отведение 1 И (3+4) 3 И 5 Пронация 1 И 5 2 и (3 + 4) Супинация 2 И (3 + 4) 1 И 5 Flexor carpi radialis Flexor carpi ulnaris Extensor carpi radialis longus » » » brevis Extensor carpi ulnaris «97 ностыо обслуживать все проходимые ими сочленения. Если максимально разогнуть кисть, то сгибатели пальцев не могут растянуться достаточно сильно; при таком положении кисти выпрямить пальцы невозможно. Наоборот, при предельно согнутой кисти невозможно крепко сжать пальцы в кулак—не могут дотянуться разгибатели пальцев. Отсюда известный прием: вынуть из ку-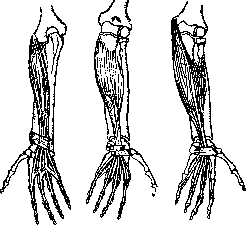
Рисунок 23. Расположзние длинных мышц пальцев. Слева Направо: extensor digitorum comm. longus, flexor dig. comm. profundus, flexor dig. comm. superficialis. Навсех рисунках ясно виден сухожильный браслет (apparatus ligamen-tosus carpi). (По МоШег'у.)
лака противника крепко зажатый им предмет, стараясь согнуть ему насильственно всю кисть. Сокращение поверхностного сгибателя пальцев вызывает движения во всех сочленениях, через которые он проходит, т. е. сгибание кисти, основных и средних фаланг пальцев, при выпрямленных ногтевых фалангах (рисунок 24, 1а). Свободно - активно (в воздухе) это движение почти невыполнимо: непременно вмешивается синергетичес-ки глубокий сгибатель, слегка огибающий и ногтевую фалангу. Вынужденно - активно это движение очень употребительно при мелких несильных движениях пальцев (движение удара на пишущей машинке, несильный фортепианный удар, держание смычка, катание пилюль, скручивание пряжи и т. п.). Движения, требующие силы и точности, всегда связаны с употреблением глубокого сгибателя, одного или совместно с поверхностным. Свободно-активное сокращение этой мышцы ведет к сгибанию кисти и сжатию пальцев в кулак (рис. 24, 2а). Это движение очень трудно выполнимо в чистом виде: привычное нам движение сжатия кулака сопряжено с разгибанием кисти, т. е. с сокращением extensores carpi (рис. 24, 2b, также lb). Эта синергия зависит от центральной нервной системы, привычно включающей всю указанную группу мышц; выше было указано, что такая синергия выгодна для силы сжатия кулака. К действиям глубокого сгибателя относятся прежде всего всякие хватки. Самые сильн. хватки объединяют все сгибатели—и короткие и длинные, включая сюда и mm. interossei, которые, не будучи в состоянии начать сгибание основных фаланг, могут однако фиксировать уже согнутые пальцы. Вынужденно-активные движения пальцев под действием глубокого сгибателя или обоих длинных сгибателей могут быть очень разнообразны в зависимости от характера воздействия внешних сил. Сюда относятся прежде всего сильные пальцевые удары (врачебная перкуссия), движения растирания (массаж, чесание), сильный нажим (прижатие струн муз. инструментов, движения лепки, формовки, врачебной пальпации и т. п.). Теми же мышцами производится далее всякая тяга с приданием пальцам формы крючка (движения пальца на эргографе, защипывание струн, поднимание тяжести за петлю и т. д.; рис. 24, За, Ъ, с). При всех этих движениях пальцы согнуты во всех сочленениях; однако если тяга за пальцевой крючок очень сильна,то палец может вынужденно выпрямиться в основной, а затем и средней фалангах, так что согнутой остается лишь ногтевая фаланга. Свободно-активно согнуть одну только концевую фалангу пальца невозможно за отсутствием подходящих мышц. Редкие субъекты способны производить такое сгибание, якобы свободно-активно, но фактически и у них оно вынужденно-активно: они всегда обладают способностью переразгибания в сочленении между основной и средней фалангой и при таком переразгибании сочленение это защелкивается у них в своего рода подвывихе. Сгибание пальцев в одних только пя-стно-фаланговых сочленениях при выпрямленных прочих (рис. 24. 4) совершается свободно-активно червячными (и межкостными) мышцами. При вынужденно-активном движении (напр. при нажиме на пистоны медных духовых инструментов, массаже, и т. п.) это движение есть уже несомненная синергия с длинными сгибателями. Разгибатель пальцев существует в единственном числе и потому выполняет все виды разгибаний; он разгибает сначала основную фалангу, а затем подтягивает остальные. Следует указать, что масса сгибателей пальцев больше массы разгибателей и пото-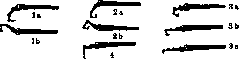
Рисунок 24. Схемы положений кисти и пальцев при различных синергиях мышц: 1 а—действие поверхностного сгибателя; lb—то же в синергии с разгибателями запястья; 2а—действие глубокого сгибателя; 2Ь—то же в синергии с разгибателями запястья; За, Ь, с—различные виды вынужденно-активных действий глубокого сгибателя; 4—действие mm. Iumbricales йшй*: et interossei.
му при спокойном положении пальцы (тонически) полусогнуты. Если разогнуть их внешней силой или собственным разгибателем, то они упруго вернутся в прежнее положение за счет тонуса сгибателей. В связи с этим надо отметить как мало известное обстоятельство,что большинство тончайших сгибательных движений пальцев совершается с помощью активного разгибания на тонусе сгибателей. Сюда относится прежде 40» всего скоропись, поскольку она не совершается сгибаниями всей кисти в запястьи. Тем же пассивным (тоническим) сгибанием после активного разгибания совершаются ажурные пассажи на фортепиано. Опытные педагоги хорошо знают это, когда тренируют па беглость гл. обр. разгибатели пальцев (упражнение с высоким подниманием пальцев). Это—один из наиболее ярких примеров использования тонуса для беглых движений.—Биомеханика мышц большого пальца вполне ясна уже из чисто анатомического их описания. 6. М. с. тазового пояса и нижней конечности. Выше было уже указано, что при переходе на прямохождение тазовому поясу пришлось приспосабливаться к новым биомеханическим условиям, совершенно отличным от тех, в к-рых он возникал и развивался. Изменение этих условий сводится к следующему. 1) На тазовой пояс легла тройная нагрузка, т. к. вес туловища, от к-рого у четвероногого 2/3 падают на передние и 1/8 на задние конечности, переложился на тазовой пояс целиком, с прибавлением еще веса верхних конечностей. 2) Клин крестца, служивший замком тазового свода и срезанный поэтому так, что его сочленовные площадки сходились к брюшной стороне, перестал отвечать своему назначению, как только весь свод таза повернулся у выпрямившегося человека почти под прямым углом к действию тяжести. 3) Затяжка symphysis pubis, лежавшая у четвероногих как-раз под крестцово-подвздошными сочленениями, в одной вертикальной плоскости с ними и с тазобедренными сочленениями, теперь оказалась впереди тех и других и поэтому перестала годиться для роли затяжки. Новой же затяжке помешало образоваться то, что на дне таза расположены выходные отверстия тела: anus, мочеиспускательный канал и, что всего важнее, родовой путь. Все особенности биомеханики человеческого таза уясняются, если понять, что он есть компромисс, труднейшая попытка возместить нарушения простой биомеханической схемы таза четвероногих, вызванные переходом на прямохождение, с помощью искривлений я переформирований частей. Человеческий таз применяется к новым механическим условиям тремя путями: 1) изменяется взаимное расположение его час-стей; 2) меняется форма костей тазового пояса; 3) усиливаются и развиваются связки в тех местах, где этого требуют усилия, возникшие в силу вертикального расположения позвоночника. Здесь уместно остановиться лишь на третьем пункте этого перечня. Т. к. затяжка symphysis pubis лежит у человека не под замком свода, а впереди от него, то развивается вторая затяжка позади свода. Эти новые затяжки соединяют tuber ischii и spina iscliii каждой стороны с крестцом (lig. tuberoso- и spinoso-sacrum), лишая при этом крестец всякой подвижности н делая его значительной помехой при родовом акте. Наряду с этими связками развиваются еще мощные связки, спирально закручивающиеся вокруг тазобедренных сочленений (lig. ilio-, ischio-, pubo-femorale). Причина их развития в том, что крестцово- подвздошные сочленения несмотря на большой компенсаторный поворот таза вперед оказываются все же лежащими на 4—5 см позади от тазобедренной оси. Поэтому туловище стремится запрокинуться относительно этой оси назад, и этому-то запрокидыванию противодействуют как упомянутые связки, так и содружественные им мышцы. Натяжение тех и других достигает при выпрямленном стоянии 20—30 кг, а при накло-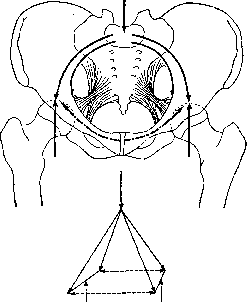
Ряс. 25. Схема статики человеческого таза. Вертикальные стрелки—нагрузка крестца от туловища и опорные реакции тазобедренных сочленений. Кривые стрелки: сплошные—линии усилий сжатия, пунктирные—линии усилий растяжения. Внизу приведена схема тех же усилий в виде палатки.
не туловища назад может достигнуть вдесятеро большего значения. Т. о. вместо ординарного кольца и ординарного свода, как у четвероногих, таз человека есть биомеханически двойное кольцо и двойной свод (рис. 25), т. е. нечто вроде палатки о четырех ножках. В силу почти полной жесткости таза как целого он почти не имеет мышечных креплений; его растяжки почти полностью связочные. Несмотря на такое резкое различие структуры плечевого и тазового поясов, еще усугубленное у прямоходящих разными условиями их механического функционирования, план мышечного оборудования того и другого сохранил единство в несравненно большей степени, нежели костный, и обнаруживает очень ясные и далеко заходящие гомологии (см. Гомологичные органы). Для их уяснения необходимо проследить прежде всего гомологию костных элементов обоих поясов, указанную в таблице. Плечевой пояс Пластина лопатки Ключица Proc. coracoid. лопатки Тазовой пояс Пластина подвздошной кости Седалищная кость Лобковая кость Эта гомология прослеживается с немень-■шей отчетливостью и на протяжении самой конечности. Верхняя конечность Нижняя конечность Шаровая головка плеча Tuberculum majus Tuberculum minus Ulna Proc. olecranon Шаровая головка бедра Trochanter major Trochanter minor Tibia Patella Эти костные гомологии в соединении с указанным выше (рис. 13) обратным поворотом передних и задних конечностей у млекопитающих помогут легко обнаружить и мышечные гомологии в обоих поясах и обеих парах конечностей. Такие гомологии изображены на рис. 26, поясняемом таблицей. По рис. ; 26 Верхняя Конечность Нижняя конечность М. latissimus dorsi М. psoas major М. teres major М. iliacus М. teres minor М. glutaeus minimus 1 4 M. infraspinatus М. glutaeus medius i 5 M. supraspinatus M. piriformis 1 _ 1 M. deltoideus (ключичная порция) M. quadratus femoris i 6 М. deltoideus (лопаточная порция) M. glutaeus maximus i 7 М. triceps brachii M. quadriceps femoris 1 8 М. biceps brachii M. biceps femoris М. pectoralis major (ключичн. порция) Mm. obturatores М. coraco-brachialis Mm. adductores femoris Механическая роль перечисленных здесь мышц в большинстве случаев вполне уясняется из их анат. расположения и из сравнения их с гомологами на верхней конечности. Особого освещения заслуживают лишь аддукторы бедра, достигающие по сравнению со своим маленьким гомологом—m. coraco-brachialis—огромного развития и включаю-
Рисунок 26. Схема гомологии мышц поясов и конечностей четвероногого млекопитающего.
щие в себя шесть анатомических наименований. По своей двигательной функции это— антагонисты для mm. glutaei medius и minimus: они двигают бедро во фронтальной плоскости в медиальном направлении. Однако это движение редкое и не требующее большой силы (т. к. совершается в направлении действия тяжести) и не может объяснить их огромного развития. Мощность этих мышц становится понятной только при оценке их статической функции. Обе ноги вместе с тазом опять-таки представляют собой арку, но лишенную уже внизу затяжки. Эту-то затяжку и заменяют аддукторы: они постоянно статически противодействуют силе тяжести туловища, стремящейся раздвинуть ноги в стороны. Гомология верхней и нижней конечностей простирается и дальше; и структура и функции mm. quadricipitis femoris точно воспроизводят таковые m. tricipitis. Однако иные условия подвижности тазобедренного сочленения позволяют отметить у этой мышцы особенность многосуставных мышц, уже отмеченную выше для длинных мышц пальцев, но не выявляющуюся у m. triceps brachii— ее недостаточную для обслуживания обоих сочленений растяжимость. Если предельно согнуть оба колена (напр. сесть на пятки), то разогнуть тазобедренные сочленения, т. е. откинуться корпусом назад, окажется уже невозможным. В еще большей степени однако это свойство выявляется у группы антагонистов m. quadricipitis, т. е. у группы сгибателей бедра и колена. Это позволяет с исключительным успехом использовать эти мышцы для измерения тонуса мышц, т. е. их пассивной сопротивляемости растяжению (Верещагин). При согнутом тазобедренном сочленении свободно-активное выпрямление колена невозможно; измерения показали, что вынужденное (пассивное) выпрямление колена в этой позе вызывает в сгибателях бедра напряжение до 70—80 кг. Это напряжение и его отношение к соответствующему процентному растяжению мышц может быть легко и точно измерено in vivo и может дать наиболее отчетливую числовую характеристику тонуса этих мышц. Ни одна другая мышца тела не может быть пассивно доведена до подобных степеней напряжения in vivo et intacto.—M. с. голени и стопы весьма проста по расположению и биомеханическому действию и не требует добавочного описания. 7. Общие условия деятельности М. с. при работе. Предшествующий функционально-анатомический обзор не может еще дать никакого представления об условиях и формах функционирования М. с. при действительной физ. работе, в реальных, а не отвлеченных условиях. Ниже будут указаны только наиболее общие, принципиальные факты, относящиеся к такой реальной жизнедеятельности М. с. Прежде всего-необходимо подчеркнуть, что деятельность М. с. в условиях работы целостного организма необычайно комплексна. При каждом двигательном (трудовом) процессе участвующие в нем так или иначе мышцы приходится считать многими десятками; при этом никогда не удается провести ясной границы между мышцами, занятыми и незанятыми в данном процессе. Если бы каждая работающая мышца обнаруживала себя напр. светящейся точкой, то ни один трудовой процесс пе имел бы не только вида звезды, но и вида целого-созвездия, а напоминал бы скорее всего млечный путь,—так обильны участвующие в процессе мышцы и так расплывчаты очертания занятых в работе мышечных участков. Выше указывалось, что напр. силовые линии m. brachialis interni прослеживаются кверху через m. deltoideus и m. trapezius до самой шеи; с еще большей сложностью сопряжены синергии при стоянии или при совместной работе рук и ног. С другой стороны существенно то, что почти никогда движение, выполняемое многоосным сочленением, не совпадает в точности по направлению с линией действия какой-либо одной мышцы, одной анат. или биомеханической единицы. Поэтому совершенно ошибочно будет представлять себе, что движение совершается за счет напряжения (или сокращения) одной мышцы при одновременном расслаблении (или растяжении) другой или других. Такое явление имеет место (и то приближенно) лишь в простейших одноосных сочленениях. В преобладающем большинстве случаев движение в данном сочленении, даже на небольшом временном отрезке этого движения, совершается за счет весьма постепенного перераспределения напряжений во всем массиве мышц, окружающих это сочленение. Если же подвергнуть рассмотрению не кусочек движения, а все б. или м. длительное движение в целом, даже движение, не меняющее от начала до конца своего направления, то окажется, что ведущая роль в движении крайне постепенно передается от одной мышцы к другой, как бы переползает по всему мышечному массиву наподобие своеобразной мышечной эстафеты.—Изучение распределения мышечных напряжений статического порядка по всему телу (что значительно проще, чем изучение динамических напряжений, и потому более далеко продвинуто) и анализ постепенных изменений этих напряжений с увеличением нагрузки и утомления приводят к ряду общих выводов, которым нельзя отказать в принципиальной важности. При статической нагрузке тела (напр. держании груза) напрягающиеся мышцы можно разделить на три разряда по фнкц. признаку. Во-первых напрягаются мышцы, деятельность к-рых непосредственно связана с удержанием груза в соответствующем положении—флексоры пальцев при держании грузов в кистях рук, бицепсы при держании на уровне груди и т. д. Во-вторых напрягаются мышцы, не влияющие сами по себе на изменение позы, не совершающие стало-быть ни одного движения при взятии груза, но потребные для фиксации поясов и туловища в условиях увеличившейся общей нагрузки. Эти мышцы осуществляют лишь повышение жесткости тех частей тела, через к-рые действие нагрузки передается на опорные части ног. Оба описанные разряда можно назвать соответственно мышцами хватки и мышцами фиксационными. Наконец в случае несимметричного положения груза (на боку или на спине и т. д.), когда его равнодействующая проходит в стороне от центра тяжести тела, напрягается еще третья функциональная группа—-мышцы уравновешивающие. При грузе на спине напрягаются таким образом mm. ilio-psoates, при держании тяжести у бока—т. erector trunci противоположной стороны и т. д. (рис. 27). Эта роль обычно выпадает на долю мощных мышц нижнего отрезка туловища. При изучении того, как один и тот же прием физ. напряжения выполняется разными испытуемыми или в разные моменты, обнаруживается, что одни из мышц обязательно участвуют в выполнении данного приема одинаково во всех случаях, другие же, напротив, то вступают в работу то бездействуют, заменяясь соседними или агонистическими, варьируя т. о. от раза к разу и от испытуемого к испытуемому. Первая категория названа облигаторными или обязательными,вторая— фоновыми мышцами (Бернштейн). Так напр.
Рисунок 27. Статические напряжения мышц при держании груза под мышкой. Возрастание густоты штриховки соответствует возрастанию степени напряжения. Незаштрихованные мышцы не обнаруживают заметной разницы с не-нагруженным стоянием.
при держании груза на носилках облигатор-но напрягаются: 1) сгибатели предплечья (кисти и пальцев), 2) m. biceps br., 3) передняя порция т. deltoidei, 4) т. rhomboideus scapulae, 5) верхняя порция т. trapezii. Легко заметить, что общие очертания всех этих мышц представляют собой своего рода лямку, перекинутую через надплечья и прямо указывающую на тот покрой, к-рый должна иметь искусственная лямка, могущая разгрузить эти мышцы. Наряду с этими «штатными» выполнителями данной работы всегда имеется налицо больший или меньший контингент внештатных фоновых мышц, обычно набираемых из числа агонистов и синергетов обязательных мышц (напр. m. brachio-radia-lis, m. brachialis int., средняя и задняя порция т. deltoidei, mm. obliqui abdominis и т. д.).—При нарастании нагрузки как правило наблюдается следующее. Для любого способа держания груза при наименьшем грузе облигаторные мышцы вступают первыми—или изолированно или в сопровождении отдельных случайных фоновых агонистов. С нарастанием нагрузки 1) возрастают напряжения в облигаторных мышцах, доходя до максимальн. величин, сопровождающихся иногда уже судорожным тремором; 2) возрастает число фоновых мышц, непо- -стоянных как по своему контингенту, так и по порядку вступления и степени нагру-женности. По мере постепенного нарастания нагрузки фоновые мышцы вербуются сперва из числа агонистов и синергетов обли-таторных мышц, а затем с нек-рого момента из числа мышц, совершенно посторонних данной статической схеме. Этот переходный момент широкой иррадиации фоновых напряжений обычно сопровождается появлением •большого количества мышц, напряженных до наивысшей степени (до тремора). Повиди-мому такой момент начала фоновой иррадиации может служить показателем предельной нагрузки, допустимой для данного ИНДИВИДУума. Н. Вернштейн. Лит.: Анатомия и гистология.-—Г е л ь в и г Р., Общая анатомия костно-мышечной системы человека, Киев, 1917; Зернов Д., Руководство описательной анатомии человека, ч. 1—Анатомия органов движения, М., 1924; I в а к и н О., До морфологии по-перекови мускулатури, Укр. мед. в1сти, 1927, № 6—7; Т о л ь д т, Анатомический атлас, вып. 3—Учение о мышцах, Петербург, 1913; Шпальтегольц В., Атлас по анатомии человека, ч. 2, М., 1907; Яцута К., Препарирование мышц, суставов и внутренностей, М., 1925: A d а с h i В., Beitrage zur Anatomie der Japaner—Statistic der Muskelvarietaten, Zeitschr. f. Morphol. imd Anthropol., B. XII, p. 261—312, 1909; Cords E., Uber funktionslose Muskeln, Ztsclir. f. d. ges. Anat., I AM., B. LXV, p. 254—276, 1922; Haeggqvist G., Muskelgewebe (Hndb. d. mikros-kop. Anatomie des Menschen, hrsg. v. W. Mollendorff, B. II, T. 2, В., печ.); Handbuch der Anatomie des Menschen, hrsg. yon K. Bardeleben, B. II—Bander, Gelenke u. Muskeln, Jena, 1904—13; Maurer F., JSntwicklung desMuskelsystems (Hndb. d. Entwicklungs-lehre, hrsg. v. O. Hertwig, B. Ill, T. 1, Jena, 1906); Mollier S., Plastische Anatomie—Die konstruktive Form des menschlichen Korpers, Miinchen, 1924; Versluys J., Das Muskelsystem (глава в книге— J. Ihle, P. van Kampen u. a., Vergleichende Anatomie •der Wirbeltiere, В., 1927). Физиология.—Б еритащвил и-Б е р и т о в И., Общая физиология мышечной и нервной системы с основами рефлексологии, Тифлис, 1927 (на груз, яз.); Бернштейн Н., Общая биомеханика, М., 1926; он же, Опыт оценки статической нагрузки мышц при держании и переносе грузов, Труды Гос. научн. ин-та охраны труда, М., 1931 (печ.); Б р у ж е с А., Метод экспериментального исследования мышечных напряжений, Исслед. Центр, ин-та труда, т. I, M., 1923; Введенский Н., О соотношениях между раздражением и возбуждением при тетанусе, СПБ, 1886; Г е р ш у н и Г., Наблюдения над прямой возбудимостью поперечнополосатой мышцы лягушки, Рус. 4>изиол. ж., 1927, № 5; Г и л л А., Работа мышц, II.—Л., 1929; Долг о-С абуров Б., Роль мышц в морфогении скелета, Изв. Ин-та имени Лесгафта, т. XVI, в. 1—2, 1930; Ефимов К., О симпатической иннервации скелетных мышц, Ирк. мед. ж., 1929, № 2; К а р ч и к я н С, Электровозбудимость нервов и мышц в агонии и после смерти, Совр. психоневр., 1928, № 7—8; КрасускаяА., Влияние деятельности мышц на их форму, группировку их волокон и способ их прикрепления, Изв. Ин-та им. Лесгафта, t,VI,1923; КрепсЕ.иСтрепьцов В., К физ.-хим. анализу симпатического влияния на скелетную мышцу, Ж. эксп. биол. и мед., 1928, № 27, стр. 558— 606; Мейергоф О., Хим. динамика жизненных явлений, М.—Л., 1926; о н ж е, Термодинамика жизненных явлений, М.—Л., 1928; Научные записки Укр. биохим. ин-та, т. V, ч. 1, Харьков, 1930 (обзор работ ин-та по биохимии мышц); Некрасов П., К вопросу о симпатической иннервации поперечнополосатой мускулатуры, Гиг. труда, 1927, № 11; П а л л а-д и н А., Хим. динамика мышц, Уеп. биол. химии, вып. 2, Л., 1925; Самойлов А., Теории мышечного сокращения, Усп. эксп. биол., т. I, в. 1, 1922; Уфлянд Ю., О мышечном тонусе, Труды Ленинградского ин-та по изуч. проф. заболеваний, т. IV, Л., 1927; Ухтомский А., Физиология двигательного аппарата, выпуск 1,Л., 1927; Чаговец В., Очерк электрических явлений на живых тканях, вып., 1, СПБ, 1903; Шохрин В., Мышечная сила в связи с возрастом, стажем и профессией, Труды Ленинградского ин-та по изуч. проф. заболеваний, т. IV, 1929; Handbuch der Biochemie des Menschen u. der Tiere, hrsg. v. C. Oppenheimer, B. IV u. VIII, Jena, 1925 (главы О. Fiirth'a, лит.); Handbuch der biol. Arbeitsmethoden, hrsg. v. E. Abderhalden, Abt. 5A — Methoden der Muskel- u. Nervenphysiologie, Berlin—Wien, с 1921; Handbuch der norm. u. pathol. Phy-siologie, herausgegeben v. A.Bethe, P. Bergmannu. a., B. VIII, T. 1—Mechanische Energie, Protoplasmabswe-gung u. Muskelphysiologie, В., 1925 (лит.); Handbuch der physiologischen Methoden, hrsg. v. R. Tigerstedt, B. II, T. 1, Lpz., 1911; Hoffmann P., Eigenreflexe menschlicher Muskeln, В., 1922; Meyerhof O., Die Energieumwandlungen im Muskel, Pflilgers Arch., Bande CLXXXII, CLXXXV, CLXXXVIII, 1920—21; он же, Die chemischen Vorgange im Muskel, В., 1930; Piper H., Elektrophysiologie menschlicher Muskeln, В., 1912; Richter F., Zur Frage nach den physikalischen Grundlagen der Muskelfunktion, Pflii-gers Arch., Band CCXVIII, H. 1, 1927; он же, Zur Theorie und Praxis der ballistischen Elastometrie, ibid., B. CCXXII, H. 1, 1928; SeilerU., Die Frage des Muskeltonus in ihrer Entwicklung u. ihrem heu-tigen Stand, Schweiz. Arch. f. Neurol., B. XVI—XVII, 1925 (также отд. изд.—Zurich, 1926); SpiegelE., Der Tonus der Skelettmuskulatur, B.,1927;Strasser H., Lehrbuch der Muskel- u. Gelenkmechanik, B. I—IV, В., 1908—17; StiibelH., Die Ursache der Doppel-brechung der quergestretften Muskelfasern, Pflugers Arch., B. CCI, 1928; V v e d e n s k у N., Du rythme musculaire dans la contraction normale, Arch, de physiologie normale et pathologique, v. Ill, 1891; Wachh older K., Beitrag zur Messung der Elasti-zitatskrafte, Pflugers Archiv, Band CCXV, Heit 4, 1926; Wright Wilhelmine, Muscle function, New York, 1928. См. также литературу к статье Животное электричество. Патология.—Б еликова О. и Смыслов Г., Фнкц. истощаемость при профессиональных заболеваниях нервной системы,Журн.невропат, и психиатр., 1926, № 6; Гутцент Ф., Подагра и ревматизм, М.—Л., 1931; Егоров Б., Болезни мышц, костей и суставов (Частная патология и терапия внутренних б-ней, под ред. Г. Ланга и Д. Плетнева, т. IV, вып. 1, М.—Л., 1928); Map сов а В., К вопросу о тонусе, Клин, мед., 1927, № 2 (обзор физиологии, патологии и проф.б-ней); о н а ж е, Метод Корнелиуса при лечении мышечных заболеваний, ibid., № 3; Резников А.,К вопросу о врожденных мышечных дефектах, Клин. ж. Сарат. ун-та, т. VII, № 3, 1929; РывлинЯ.,К вопросу о профессиональных разрывах мышц, Нов. хир., 1928, № 10, стр. 544; Хорош-к о В., О нервно-мышечном перетруживании, Рус. клин., 1927, № 3; A b r i k о s s о f f A., Uber Myome, ausgehend von der quergestreiften willkurlichen Mus-kulatur, Virchows Arch., B. CCLX, 1926; В e n e к e, Uber Muskelveranderung bei Intoxikation und Infek-tion, Verhandl. der deutsch. pathol. Gesellschaft, Berlin, 1913; Durante G-., Anatomie pathologique des muscles (Manuel d'histologie pathologique, sous la dir. de Cornil et Ranvier, v. II, P., 1902); Handbuch der speziellen pathologischen Anatomie u. Histologie, hrsg. v. F. Henke u. O. Lubarsch, B. IX—Knochen u. Muskeln, T. 1—2, В., 1929; Kttttner H.u.Lan-d о i s F., Die Chirurgie der quergestreiften Musku-latur, Stuttgart, 1913; Lindstedt F., Uber die Natur der muskelrheumatischen (myalgischen) Schmerz-symptome, Acta med. Scandinav., supp. 30, p. 1—180, 1929 (исчерпывающ, лит.); L о m m e 1 F., Erkrankun-gen der Muskeln, Knochen u. Gelenke (Hndb. d. inn. Med., hrsg. v. G. Bergmann u. R. Staehelin, B. IV, T. 1, В., 1926; L о r e n z H., Die Muskelerkrankungen, Wien, 1904; Marinesco, Maladies des muscles (Nouveau traite de medicine, sous la dir. de A. Gilbert et P. Carnot,'v. XXXVIII, P., 1910); T e s t u t J., Les anomalies des muscles chez 1'homme, P., 1884; T h i e г s J., Affections des muscles (Nouveau traite de medecine, sous la dir. de H. Roger, F. Widal et P. Teissier, v. XXII, P., 1924); T h о m s о n F. a. G о r d о n R., Chronic rheumatic diseases, London 1926 (русское издание—Москва-Ленинград, 1928); W e r t h e i m-S a-lomonson J., Myositis Handbuch der Neurologie, herausgegeben von M. Lewandowsky, Band II, Berlin, 1911, литература). Смотрите также:
Смотрите также:
- МЫШИ, мелкие грызуны, образующие вместе с крысами подсемейство Murinae.— Mus musculus, домовая мышь, является космополитом, распространившимся с человеком по всему свету. Живет в домах и службах; может обитать в садах ...
- МЫШИНЫЙ ТИФ, б-нь, впервые описанная Лефлером (1892) как эпизоотия среди белых лабораторных мышей в Грейфсвальде. Лефлер определил возбудителя этой эпизоотии в виде бактерии, сходной с бактерией брюшного тифа, почему и назвал ...
- МЫШЛЕНИЕ, в психологии—высшая и наиболее сложная форма интелектуальной деятельности, состоящая в рациональной переработке данных опыта, в процессах установления связей, вскрытия отношений и зависимостей и отличающаяся своеобразным составом, строением и способом ...
- МЫШЦЫ. I. Гистология. Общеморфодогически ткань сократительного вещества характеризуется наличием диференцировки в протоплазме ее элементов специфич. фибрилярной структуры; последние пространственно ориентированы в направлении их сокращения и являются мор-фол. субстратом специф. сократит, ...
- МЫШЦЫ ЧЕЛОВЕКА ...