РЕГЕНЕРАЦИЯ
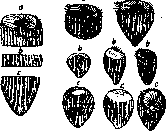
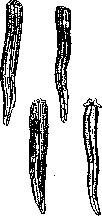

Рисунок 2. Регенерация морской звезды Linckia mul-
tifofa из одного луча. Последовательные стадии регенерации. (По Коршельту.) и кожные покровы. Наконец млекопитающие и в том числе человек способны к замещению лишь небольших участков органов и кожных повреждений. Регенерационная способность не остается одинаково выраженной в течение всей жизни индивидуума: различные стадии.развития отличаются в этом отношении каждая своими характерными особенностями. Как правило можно сказать, что, чем моложе животное, тем выше его регенерационная способность. Головастик например может на ранних стадиях развития регенерировать конечности, в то время как, вступая в период метаморфоза, он эту способность теряет. Указанное общее правило имеет однако ряд исключений. Известны случаи, когда более ранний стадий развития обладает меньшей регенерационной способностью. Личинки планарий отличаются меньшим разви- 685 РЕГЕНЕРАЦИЯ 536 тием ре генерационных явлений по сравнению со взрослыми животными (Steinmann), то же имеет место для личинок нек-рых других животных. Уже из вышесказанного можно было видеть, что различные области организма отличаются друг от друга по своей регенерационной способности. Вейсман принимал, что способ- ность Р. зависит Рн'и{ [ [ | | | ( [ | от того, насколько данная -часть подвержена опае-. ности повреждения, причем чем больше последняя, тем больше и регенерацион-ная способность ,— свойство, выработанное в результате естественного отбора. Однако поел едние исследования показали, что такая закономерность не 6,6 15 6,9 10 7,2 5 ■ ■\ г°\ /i [^ 1 * .у/ ""ч> *■-.„ 8 Ю 12 14 Рисунок 3. Сплошная линия—изменение интенсивности митогене-тического излучения регенерирующего хвоста аксолотля. Н? ордин .те условные единицы интенсивности излучения. Прерывистая линия — изменения активной реакциитканей регенерирующей конечности аксолотля. На ординате—значения рН (дан- может быть уста-ные Окунева). На. абсциссе—дни НГШ7ТРТТЯ. ^„^ пл-регенерации. (Из Бляхера и ноялена. ряд ор Бромлей.) ганов, не подвер- гающихся обычно повреждению в течение свободной жизни индивидуума и хорошо защищенных, обладает тем не менее высокой регенерационной способностью (Morgan, Przibfam). Убиш (Ubisch) связывает регенерационные явления с диференци-ровкой организма; по его мнению ранее развивающиеся части скорее прекращают регенерировать с возрастом или их Р. отличается меньшей интенсивностью. Так, у амфибий, где диференцируются раньше органы, лежащие более кпереди, можно установить соответствующий градиент Р.—спереди назад. Утверждения Убиша, в пользу к-рых говорит ряд данных, нуждаются все же в дальнейшем подтверждении на большем материале. На некоторых видах (преимущественно на червях) Чайлд (Child) и его сотрудники установили точно так же определенный градиент Р. по отношению к продольной оси тела, но направление его не всегда идет спереди назад, а связано с более сложными закономерностями. Чайлд считает, что этот градиент зависит от степени физиол. активности различных участков организма. Более низко организованные животные обладают способностью регенерировать как части, расположенные проксимально от места ампутации, так и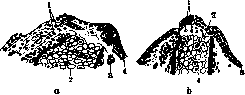
Рис 4. Регенерация ампутированной передней конечности у саламандры через */4 (а) и 12 (Ь) часов, a: i—бластемные клетки; 2—культя плеча; 3—нерв; 4—эпидермис; Ь: 1—бластемные клетки; 2—хрящ; 3—эпидермис; 4—культя плеча.
расположенные дистально. У высших животных регенерируют лишь последние.У амфибии напр. орган, даже пересаженный в перевернутом положении, регенерирует то же образование, что и в обычном положении.
Рисунок 5.:Регенерап*яг «ам-
Течение регенерационного процесса. Регенерационный процесс протекает различно в зависимости от того, с каким организмом'мы имеем дело и какая часть его подвергается удалению. В качестве примера можно рассмотреть наиболее изученный объект— Р. конечности амфибий. При этом имеют место следующие явления. После ампутации органа происходит сближение краев раны вследствие сокращения перерезанной мускулатуры. Находящаяся наповерхно-сти раны кровь свертывается, выделяя нити фибрина. Свернувшаяся путированной"''передней Кровь при участии ПО- конечности у саламандры ттржттрнтттлтг ткянрй пб- чрррз 8 дней: J и 2-бла" врежденных тканей оо- стемные клетки; з—эпи- разует на раневой ПО- дермис; 4—культя плеча. верхностй струп. В результате повреждения тканей и воздействия внешней среды на незащищенную кожей поверхность в органе возникают процессы распада. Последние выявляются в изменении кислотности регенерата (понижение рН от 7,2 до 6,8, Окунев) и появлении митогенетического излучения (Бляхер и Бромлей). Раневая поверхность не остается однако долго незащищенной: уже в течение ближайших часов наблюдается процесс наползания эпителия с краев раны, в результате чего на раневой поверхности образуется эпителиальная пленка. Под этим эпителиальным покровом происходятвседальнейшие процессы, сводящиеся к разруше- нию и перестройке старого и образованию нового органа. Эти про-?VІ^*i цессы выражают-л° ' ся, с одной стороны, в продолжающемся распаде. Последний выяв- Рисунок G. Регенерация передней лен морфологиче-конечности у саламандры через СКИ В резуЛЬТа-9 дней: 1—гигантские клетки; тр гигт иррирттп-2—бластемные клетки; л—нудь- Те ГИСТ" исолеЛи тя плеча; 4—мускулатура; 5— Вания, ПОКаЗЫВа-эпидермис. ющего ' картины разрушения тканей и прихода в регенерат многочисленных кровяных клеток. Распад особенно силен в период от 5 до 10 дней, начиная с момента ампутации, когда он достигает повидимому наибольшей интенсивности. Об этом же свидетельствуют и физиологические показатели. Окунев* нашел наибольшую кислотность на 5-й день, когда рН=6,6. Усиливается одновременно и интенсивность митогенетического излучения по сравнению с предыдущими днями (Бромлей). Кривые повышения кислотности и интенсивности митогенетического излучения оказываются параллельными друг другу на всем протяжении регенерации. И та и другая ■ имеют две вершины максимума—на 1-й и 5-й день Р. (рис. 3). Наряду с этим уже на первой неделе Р. ясно обозначаются новообразовательные процессы. Они сказываются преимущественно в образовании под эпителиальной пленкой разрастания из однородных клеток, носящего название бластемы. Развитие нового органа идет преимуще-
Рисунок 7. Регенерация ам-
■ственно за счет клеток бластемы (рис. 4—7). После известного периода роста в регенерате происходит диференциация отдельных частей. При .этом сначала диференцируются более проксимальные части, а затем уже дистальные. В этом •отношении не у всех организмов процесс течет одинаково. У некоторых _^Щ|^ животных отношения ^ШШЁ!%ч даже могут быть обрат- ными, Физиол. особенности регенерата конеч--2 но не те, что у сформированного органа. Это проявляется в частности 11 в том, что регенерат об-j ладает гистолизирую-^ щими свойствами. В том случае, когда поверхность его приходит в соприкосновение с другими тканями, напр. при закрытиирегенерата ко- путированной "передней ЖНЫМ ЛОСКутОМ, насту-конечносты у саламандры пает ГИСТОЛИЗ ПОСЛвД- стеРм^\Теетки/2"-ги: них (Бромлей и Орехо-гантские клетки; з—эпи- вич). Не следует думать, дермис; 4—мускулатура; что процесс Р. СКаэыва-5—плечевое кольцо; 6'— р _ тгпькп на амггети-культя плеча. (По Кор- ется только Hcl амиуш шельту.) рованном, регенерирую- щем органе. Он оказывает свое воздействие и на остальной организм, что может проявляться в различных отношениях. Так, изменение может быть уловлено в крови животного, митогенетическое излучение которой отклоняется от нормальной интенсивности, причем" колебания эти имеют характерную кривую. При Р. у гидр отмечен распад органов, не находящихся в непосредственной •близости с регенератом,именно половых клеток, притом преимущественно мужских (Goetsch). Влияние Р. сказывается также на росте и иных свойствах организма — явление, б. ч. •описываемое под названием регуляции. Материал регенерата. Вопрос о материале, за счет к-рого происходит образование регенерата, должен разрешаться различно в зависимости от вида животного и характера нанесенного повреждения. Если дело идет о повреждении одной какой-либо ткани, то обычно процесс идет за счет разрастания остатка •соответствующей ткани. Сложнее обстоит дело в случае Р. органа или восстановления организма из отдельного участка его. При этом однако можно установить, что в основном, по крайней мере у амфибий, Р. идет за счет материала, непосредственно прилежащего к раневой поверхности, а не за счет клеток, приходящих из других областей организма. Это показывают олыты Р. гаплоидноядерноЙ конечности тритона, пересаженной на диплоидноядер-ное животное. Возникающий при этом регенерат состоит из гаплоид ноя дер ных клеток (Hert-wig). To же следует из пересадок конечностей от черной расы аксолотлей к белой, когда регенерирующая конечность оказывается черной. Факты этл исключают представление о Р. за счет различных клеточных элементов, приходящих с током крови. При рассмотрении материала, идущего на Р., приходится считаться с двоякой возможностью. Р. может происходить или за счет так наз. резервных, индиферент-ных-клеток, остающихся недиференцированны-ми во время эмбрионального развития, или же имеет место использование уже специализиро-
вавшихся клеточных элементов. Важное значение резервных клеток было показано для ряда животных. Так, Р. у гидр происходит в основном за счет т. н. интерстициальных клеток. То же имеет место у турбеллярий. У кольчецов эта роль принадлежит необластам, относящимся к подобного же рода элементам. У ас-цидий индиферентные клетки также играют важную роль в Р. Сложнее обстоит дело у позвоночных, где различные авторы приписывают основную роль в Р. разным тканям. Хотя и здесь имеются указания на происхождение клеток бластемы из неспециализированных элементов, однако факт этот нельзя считать прочно установленным. Тем не менее положения господствовавшей ранее теории Gewebe-sprossung, признававшей возможность развития клеток какой-либо ткани лишь из клеток подобной же ткани, были основательно поколеблены. Но если можно принять образование значительной массы регенерата за счет неспециализированных клеток, то это не исключает и возможности развития части регенерата из дифе-ренцированных элементов. При этом может итти речь как о развитии тканей -за счет размножения одноименных элементов, так и о переходе клеток одного типа в другой (метаплазия). На самом деле во многих случаях можно показать, что имеют место оба-эти процесса. Так, мускулатура обычно В значительной Рисунок 8. Рентгенограм-чаСТИ ПРОИСХОДИТ ИЗ ОСТав- ма регенерации бес-шихся неразрушенными $ЖсЖ™?^ мышечных клеток. У кольчецов можно установить образование мышц из эпителиальных элементов. То же имеет место у нек-рых раков (Пржибрам). Образование нервной системы из эктодермальных клеток установлено у асцидий (Schultze). У амфибий известно, что Р. линзы может происходить/из края радужины (Wolff, Colucci). Также можно принять образование хрящевого и костного скелета без участия хрящевых -и костных элементов ранее существовавшего органа.
Поскольку регенерационный процесс включает в себя как. развитие из индиферентных элементов, так и участие специализированных элементов, то в каждом отдельном случае необходимо специальное исследование для выяснения роли каждого из,этих процессов в Р. Если рассмотреть как пример Р. у амфибий, опять-таки в силу ее наибольшей изученности, то дело здесь представляется в следующем виде. Нервы всегда образуются за счет роста окончаний старых нервных стволов. Иначе обстоит дело с костной тканью в случае Р. конечности. Было показано, что даже при удалении всего костного скелета конечности, включая и плечевой пояс, при ампутации такой бескостной конечности наступает Р. органа, обладающего скелетом (Fritsch, 1911; Weiss, Bischler) (рис. 8). Иначе обстоит дело при Р. хвоста. В этом случае костные части образуются лишь тогда, когда имеется в области регенерата повреждение старых скелетных частей, лены плечевой пояс и плечо; ампутация выше локтя. Регенерированы предплечье с костями предплечья и кисть с фалангами. Carpus еще хрящевой, radius и ulna сдвинуты в бескостное плечо. (По Кор-шельту.)
н костные элементы последнего могут принимать участие в Р. (рис, 9). Относительно соединительнотканной части кожи, corium, мы также имеем доказательство возможности ее образования без участия старого corium а (Вейс), Что касается мускулатуры, то удаление большей части мускулатуры конечности не приводило к каким-либо аномалиям в развитии регенерата. Кроме того в случае пересадки кусочка хорды у личинки Anura в область хвоста, лишенную мускулатуры, удавалось вызвать образование хвоста в этом месте при соотв. направлении разреза хвоста. Образующийся при этом орган обладал мускулатурой (Marcucci). Однако гистологические исследования показывают, что при обычной Р. хвоста мышцы его образуются из соответствующих же элементов старого органа (NaVІlle). Так. обр. значительная часть регенерата у амфибий *может образоваться не в результате размножения старых тканей, а из массы бластемы, происхождение элементов которой, как уже указывалось, не установлено еще в достаточной мере. В то же время могут иметь место и иные отношения, что мы имеем при Р. хвоста, осевые органы й-рого регенерируют лишь при наличии старых. При этом следует отметить, что даже Р. одного и того же органа Может итти за счет различного материала в зависимости от условий, как можно было убедиться на примере образования мышечных элементов хвоста. Приведенные опыты, хотя и указывают на возможность развития нек-рых тканей (напр. костной) не из клеток подобной же ткани, не разрешают все же вопроса о том, как обстоит дело при нормальных условиях Р. В этом направлении необходимы дальнейшие исследования.
Условия Р. А. Регенерирующая область. Течение Р. конечно находится в тесной зависимости от того, какой участок организма подвергается ампутации и следовательно в какой области разыгрываются регенераци-онные явления. Прежде всего мыможем столкнуться с отсутствием Р. в нек-рых частях организма или вернее со слабым выражением соответствующих явлений. Филиппо (Philippeau) обнаружил отсутствие регенерации у саламандры в случае экстирпации конечности со всем плечевым поясом. Шотте (Schotte) показал, что ампутация хвоста сопровождается регенерацией лишь в том Рисунок 9. Рентгенограмма регенерировавшего хвоста ящерицы Lacerta mu-ralis. Разрыв вобласти IV хвостовэго позвонка.(По Коршельту.)
Рисунок 10. Triton cristatns после полного удаления территории хвоста; никаких следов регенерации в течение 8 месяцев.
случае, если разрез проходит достаточно ди-стально (рис. 10). Валлет и Гиено (Vallette, Guyenot) отмечают отсутствие регенерации носовых частей головы при ампутации слишком большого участка. Точно так же Р.,глаза не происходит при полной энуклеации (Шак-сель). Жабры при полном удалении не регенерируют. Гиено толкует эти явления таким образом, что Р. может происходить лишь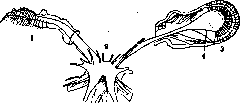
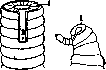
Рисунок 12. Регенерация переднего отдела у дождевого червя. Положение регенерата определяется нервным стволом: 1— плоскость регенерации; 2—конец перерезанного нервного ствола.
Рисунок 11. Замена левого глаза, удаленного вместе с глазным ганглием, антеннообразным придатком (I): 2—надглоточный ганглий; 3 — глаз; 4—глазной ганглий. (По Коршельту.) при наличии определенных клеточных комплексов, к-рые могут быть полностью удалены при достаточной степени повреждения. Достоверное доказательство этого положения однако пока еще не дано, и не исключено, что в некоторых случаях отсутствие регенерации, обнаруженное указанными авторами, связано и с иными условиями. От регенерирующего участка зависит также характер того образования, которое возникает при Р. Хорошо известно, что при удалении различных частей организма возникают различные же образования. Не следует однако объяснять это явление тем, что новообразующийся орган должен быть сходен с удаленным. Так, известен опыт Гербста (Herbst), подтвержденный и другими авторами, когда при удалении у рака глаза при оставлении зрительного ганглия регенерирует глаз, а при одновременном удалении и ганглия наблюдается Р. антенны (рис. 11). При экстирпации у одного вида насекомых (Dixippus morosus) усика в дистальной части наблюдается образование усика, при ампутации же у основания регенерирует конечность. Соответствующие явления носят название гомойозиса. Понятно, что от регенерирующего участка зависит также и скорость Р., о чем уже говорилось. Б. Части ампутированного органа. Как видно было из опытов удаления скелета конечности, Р. может иметь место и в его отсутствии. Однако, как показала Бишлер,. при Р. бескостного органа регенерирует не тог сегмент, к-рый подвергается ампутации, а лишь более дистальный, так что при Р. напр. конечности возникает орган, укороченный на один сегмент. Поскольку развитие наблюдается и в отсутствии костной ткани, то связь специфичности Р. со скелетом отрицается. Кроме того> пересадки одних костей на место других, напр. бедра на место плеча,не изменяют морфологии регенерата. Важная роль в регенерационных явлениях принадлежит нервной системе. Необходимость наличия нервных связей для образования регенерата доказана, однако не для всех видов. Для ряда животных такой законо- 54£
мерности повидимому не существует. Наиболее ясные данные имеются по червям, иглокожим и особенно амфибиям.. У червей Морган показал необходимость наличия нервных окончаний в подвергающемся Р. участке для того, чтобы регенерационный процесс мог иметь место (рис. 12). То же показано и для морских звезд (Мог-gulis). Однако имеются данные, противоречащие толь-ко-что упомянутым, так что в этом йаправлении необходимы дальнейшие исследования. Для амфибий показано, что наличие центральной нервной системы не является необходимым условием P. (Barfurth, Рубин, Годлевский). Однако в случае нарушения периферической иннервации Рисунок 13. Гетеротопи-регенерирующего органа ™чЯенная°путемСот1 процесс восстановления ОТ- ведения плечевого СуТСТВует. Имеющие здесь сплетения. (По Гие-место отношения были вы- н0-)
яснены в результате подробных опытов Шотте и Вейса. Оба они показали, что в случае полной денервации Р. не имеет места. Шотте показал, что при этом имеет значение лишь симпат. нервная система, т. к. при перерезке симпат. нервов и оставлении чувствительной и моторной иннервации образование органа не происходит. Наоборот, Р. налицо при сохранении одной симпат. иннервации. Значение нервной системы доказано Шотте не только для взрослых животных, но и для личинок. Данные Шотте в отно-: шении симпат. иннервации однако вызывают возражение у некоторых авторов, считающих, что основная роль в регенерационном процессе принадлежит спинальным ганглиям (Locatelli). Полученные данные говорят также о том, что роль нервной системы не ограничивается только начальными стадиями процесса; для продолжения Р. наличие нервной системы также является ; необходимым. Ряд авторов ставит специфичность регенерата в связь с нервной системой. По их мнению имеется специфическое влияние последней. Интересные данные в пользу этого предположения были приведены Локателли, которая получала образование добавочных конечностей у тритонов путем выведения центрального конца перерезанного п. ischiadici на поверхность тела в области бока и задней конечности (рисунок 13). Однако Гиено и Шотте показали своими исследования- ; Жк, что специфичность нервов не играет роли в данном явлении. Правда, приведение перерезанного конца нерва в ту или иную область Организма вызывает здесь образование органа, однако характер органа связан со специфичностью области, а не нерва. Один и тот же нерв, будучи приведен в участок, окружающий заднюю конечность, вызывает здесь развитие зад-! ней ноги, а попадая в участок, расположенный ближе к хвосту, вызывает образование, именно последнего органа. При приведении нерва в промежуточные области можно полу-
Рисунок 14. Заторможенная регенерация правой задней конечности аксолотля вследствие образования рубца кожи. (По Кор-шельту.)
чить химерические образования между хвостом и конечностью. Ряд других данных в пользу специфичности нервной системы (Вольф, Walter) также получил иное объяснение. В связи с этим предположение о специфичности нервного влияния на Р. должно быть отвергнуто. Удаление кожи в месте ампутации на известном протяжении приводит к тому, что Р.. органа задерживается до тех пор, пока эпителий, наползая с края кожи на открытую поверхность, не покроет ее и не дойдет до места ампутации. Может наступить также дегенерация открытого участка и тогда Р. начинается с того момента, когда дегенерация участка дойдет до края кожи и соответствующие части отвалятся. Т. о. наличие кожи, вернее эпителиального покрова, является необходимым условием Р. органа. Положение это объясняет отсутствие Р. при закрытии раневой поверхности кожным лоскутом (рис. 14), показанное рядом авторов как на амфибиях (Tornier, Шаксель, Годлевский, Ефимов), так и на насекомых (Шаксель и Adensamer). Явление это обусловливается тем, что эпителий кожи не имеет доступа к раневой поверхности, будучи отделен от нее соединительнотканной частью кожи, для наличия же Р. необходимо покрытие рацы молодым эпителием. Если под лоскут кожи,. покрывающий раневую поверхность, пересадить кусочек кожи, то Р. в этих случаях наступает (Ефимов). Факт этот говорит за то, что механическое препятствие для роста регенерата в этом явлении не играет роли. Специфичность кожи не влияет на характер регенерата. За это говорит опыт Таубе, пересаживавшего манжетку красной кожи живота, у тритонов на конечность и получившего после Р. из места, покрытого красной кожей, обычную черную конечность. То же подтверждает и пересадка внутренних частей хвоста в кожный рукав конечности, когда наблюдается у. хвоста (Бишлер). Удаление большей части Мускулатуры оказывает влияние лишь на скорость процесса. Приходится отрицать также и специфическое влияние мускулатуры, поскольку замена путем пересадки мускулатуры одной области на иную не изменяет характера регенерата (Бишлер). Приходится т. о. признать, что каждая из упомянутых частей. органа (нервы, скелет, мускулатура, кожа), взятая в отдельности, не является специфическим условием Р. В. Части регенерата. Регенерирующий орган неоднороден не только в том смысле г что состоит из разных тканей, в нем имеются участки, чрезвычайно сильно отличающиеся друг от друга по своим свойствам. Если разделить регенерирующий орган на две различные части, как это обычно делается, бластему и остальной регенерат, то поведение их оказывается резко отличным. При удалении бластемы последняя вновь образуется остающимися частями, то же происходит при пересадке части органа, не заключающего бластему, в какую-нибудь иную область организма. При этом даже очень небольшие кусочки трансплянтированно-го участка могут развить соответствующий орган (рис. 15). Иначе обстоит дело при пересадке другой части регенерата—бластемы. При этом обнаружилось, что до известного возраста, примерно двух недель, бластемы, будучи пересажены, не развиваются дальше и рассасываются (Шаксель). Бластемы в опытах де Джорджи (de Giorgi), пересаженные на спину в воз- ■513
Рисунок 15. Результаты ■трансплантации тер-
неи конечности на место хвоста. (По Гие-но и Понс.) расте до 30 дней, хотя и приживлялись и несколько увеличивались, но диференцировки не испытывали. Какого рода условия имеют тут .значение, сказать трудно, во всяком случае вывод из указанных фактов может быть лишь тот, что для наличия Р. необходима связь между бластемой и остальными частями регенерата. Ряд авторов пытался выяснить, какая .именно часть регенерирующего органа является специфичной,отличающей один орган от другого. Особенно много внимания было уделено вопросу о том, является ли специфичным материал бластемы. Соответствующие исследования сводились к пересадкам бластем одних органов на другие с целью выяснения, изменится ли при этом специфичность образующегося из бластемы органа. Пересадки бластем производились на различных видах животных. При этом обнаружилось, что регенерат, пересаженный до известного возраста, развивается в соответ-ритории хвоста на ме- rrpRWw г той пбняотьто rm-сто плеча и фрагмен- ствии с тои ООЛастью орта территории перед- ганизма, на которую он пересаживается. Т. о. эти опыты говорят за неспецифичность бластем. Однако все до сих пор проведенные исследования не являются достаточно убедительными. Милоевич •(MiloseVІc) при пересадке молодых регенератов задней конечности на место передней получил в ряде случаев образование на новом месте передней конечности, т. е. развитие сообразно месту пересадки. Однако данные эти не доказательны вследствие отсутствия достоверного критерия того, что образующийся орган происходит действительно из ткани тран-сплянтата, а не самой регенерирующей передней конечности. В опыте Гиено и Шотте, где бластема конечности, будучи пересажена на хвост, дала образование хвоста, сами авторы сомневаются в происхождении материала органа: Наконец Вейс пересаживал регенераты хвоста в область передней конечности и получил в трех случаях развитие конечности. Однако и в этих опытах не может быть уверенности относительно того, за счет ли тканей трансплянтата идет Р. Таким образом вопрос о возможности изменения пути развития регенерата у амфибий, а вместе с тем и вопрос о специфичности бластемы, остается открытым. Аналогичное положение имеет место и для низших животных. Опыты Гебгардта, получившего в двух случаях образование головы из регенерацион-ной почки хвоста у планарии, могут быть истолкованы как результат участия в регенерации тканей головной области, куда производилась пересадка. Все сказанное касается только молодых регенератов, поскольку все авторы сходятся в том, что новообразующиеся ткани, взятые в относительно позднем возрасте, отличаются уже специфичностью. Несмотря на недостаточную очевидность опытов пересадки молодых регенератов большинство авторов считает специфичной не бластему, а лишь остальную часть органа. Наличие мито-тенетического излучения, в регенерате позво- лило высказать мысль о возможности влияния ■ излучения одних частей регенерата на другие, особенно митогенетических лучей, возникающих при резорпции тканей, на размножение клеток бластемы (Бляхер и Бромлей). Пока еще однако значение митогенетического излучения при Р. не может считаться установленным. Несомненно все же, что, воздействуя ми-то генетическими лучами на регенерат, мбжно вызвать ускорение процесса (Бляхер, Воронцова, Ирихимович, Лиознер). Те же авторы показали наличие стимуляции регенерационных процессов в тех случаях, когда раневые поверхности имеют возможность влиять друг на друга (например при треугольном вырезе участка хвоста). Г. Процессы, происходящие в организме во времярегенер а ц и и. Р. является процессом, зависящим не только от состояния данного органа, но и от всего организма. Поэтому процессы, происходящие в последнем, могут иметь решающее влияние на регенерационный процесс. В опытах Геча ампутация головы у гидры не вела к Р. в том случае, когда гидра обладала почкой. Тогда происходили лишь регулятивные процессы, в результате которых голова увеличивающейся почки занимает место головы полипа. Если у двухголовой планарии ампутировать одну голову, то последняя не регенерирует (Штейн-ман). Изменение локализации регенерирующего органа по отношению к организму может не оказать, однако влияния на характер регенерации. Курц (Kurz) пересаживал ампутированную конечность на спину, причем здесь регенерировала нормальная конечность. Вейс менял местами передние и задние конечности тритона и опять-таки Р. пересаженных конечностей приводила к развитию того органа, который образовался бы в случае оставления их на месте. То же имеет место при пересадке участка хвоста или передней части головы. Т. о. то или иное место развития процесса не является специфичным при Р. Влияние организма на Р. его частей может сказаться не только на обусловливании самой возможности Р., но и на характере регенерата, его форме, положении и течении процесса. Примером такого воздействия может служить например значение функции для регенерационного процесса, когда употребление органа сильно сказывается на регенерате. Значение других частей организма для Р. данной области выявляется в опытах с инкреторными железами; удаление желез внутренней секреции или воздействие их инкретами может оказать влияние на ход Р. Несомненно, что целый ряд процессов, происходящих в организме, оказывает воздействие на регенерационный процесс. Из них можно упомянуть случаи одновременно наличия в организме нескольких регенерационных процессов. Произойдет ли при этом стимуляция или торможение Р.—зависит от конкретных условий, выражающихся в размере этих повреждений, расположении их и т. д. (Zeleny). Влияние имеющихся в организме связей на Р. сказывается в опытах вырезания из тела гидр или планарии небольших участков. При этом может наступить извращение полярности, когда на обеих сторонах регенерата образуются одинаковые органы (образование животных с двумя головами или двумя хвостами в зависимости от той области, из которой вырезался регенерирующий участок).
Д. Окружающая среда. Что Р. может протекать лишь в соответствующей среде, достаточно очевидно. При составе среды, вредно действующем на ткани, регенерационный процесс конечно невозможен. Для нормального течения Р. окружающая среда должна отвечать ряду условий. К ним относится прежде всего определенное содержание кислорода (Леб). Далее Р. возможна лишь в определенных температурных границах. Оптимум для амфибий напр. равен 28°, выше и ниже этой температуры Р. замедляется, при 10° она совсем прекращается. По исследованию Мура (Мооге) скорость Р. в зависимости от t° подчиняется закону вант Гоффа. Для водных животных большое значение имеет состав окружающей их жидкости. Р. возможна лишь при определенной концентрации морской воды (Леб, Штейнман). Наилучшая Р. наблюдается в разведенной морской воде. Нек-рые соли (калий, магний) оказываются также необходимыми для наличия регенерационного про-
Рис 16. Хвостовые Чесса (Леб)> ДРУгие ока-отрезки Pianaria go- зывают влияние на ско-nocephala с регенера- рость его. Попов получил при воздействии1; рад значительную стимуляцию ь—без воздействия; регенерационного процес-A~B??5/eEMBpie 5 т^Г са' В03ДеиствУя на плана- в^действие 10 минут" Рий и ПОЛИПОВ раСТВОра- танином+KJ—через4 ми MgCl2, KJ с глицери- дня; С-то же через 7 ном, танином и др. веще- дней.(ПоКоршельту.) ствами (рисунок 16). Сти. мулирующее действие на регенерацию оказывают также вещества, понижающие поверхностное натяжение среды, Е. Характер повреждения. Регенерационный процесс зависит не только от той области, где производится ампутация, но и от характера повреждения. При небольшом порезе на стенке тела животного может наступить быстрое заживление при почти полном отсутствии новообразования тканей. При нанесении однако в этом же месте нескольких насечек, мешающих такому заживлению,на- ^ .„ „ г vrmnmn Рис' 17' Развитие гидранта из ^xyiicici лириши боковой области полипа Corymor-выраженный ре- pha palma под влиянием радиаль-генерационный ных надрезов: I—надрезы; 2, 3, ттпттарр r пр 4—постепенное развитие гидран-процесс, в ре- та. (Из Чайлда.) зультате к-рого развивается целый орган (напр. голова животного; Леб, Чайлд) (рис. 17). От характера повреждения может зависеть атипичное течение Р. Так, при раздвоении ампутируемого органа возникают двойные образования. Положение регенерата также может зависеть от того, как производится ампутация, поскольку длинная ось возникающего регенерата обычно перпендикулярна к плоскости ампутации. Теории Р. Явление Р. стало известно очень давно. У ряда ученых древнего времени можно найти указания на знакомство с этим явлением. Однако систематические опыты, посвященные изучению Р., были поставлены уже ближе к современности. Реомюр (Reaumure) изучил регенерацию у рака, приписывая это явление наличию добавочных "зачатков органов (1721). Известны данные Тремблея на гидрах, относящиеся к 1744 году, установившие отчетливо выраженную регенерационную способность этого животного. Середина и конец 18 в. насчитывают еще ряд исследований по Р. Сюда относятся данные Бонне и Спалланцани (Bonnet, Spallanzani). Исследования' эти захватывают не только низших, но и ряд высших животных (позвоночные). В ближайшие затем годы изучение Р. продвигалось очень медленно. Лишь в конце 19 века начинается усиленное исследование регенерационных явлений, охватывающее самые различные типы животных. Изучение это характерно не только своей систематичностью и детальностью, но и тем, что исследователи уже значительно глубже проникают в сущность явления Р. Исследователи конца 19 в. много внимания уделяют выяснению связей регенерационного процесса, его необходимых условий и на этом материале строят соответствующие теории Р. Принципиальный подход этих авторов к изучению процесса получил свое обоснование в работах В. Ру и может быть назван каузально-аналитическим методом исследования. Характерными его чертами являются механистичность и формальность анализа явлений; моменты, приводящие к возникновению изучаемого явления, берутся не в процессе развития, а как неподвижные. Разлагая процесс на отдельные компоненты, выделяют основной компонент,, который принимается за исходное, и само явление рассматривается как результат воздействия на эту основу различных условий. С другой стороны, т.к. направление процесса рассматривается изолированно от его движущих сил, то выделяется также на основании формального анализа отдельный фактор, ответственный за направление процесса. Т. о. источники развития и направления явления оказываются внешними по отношению к отдельным компонентам процесса. Поскольку источник развития выступает как внешний по отношению к остальным компонентам процесса, то неизбежен вопрос о том, чем вызывается развитие самого источника развития. Если в качестве последнего будет выделен какой-либо фактор, то вновь встанет вопрос об источнике развития этого нового фактора. Идя т. о., мы или должны притти к божественному первотолчку или ртказаться от окончательного разрешения вопроса. Вся неправильность каузально-аналитического метода ясно вытекает уже из этого его описания. Общность метода не мешает однако исследователям Р. расходиться между собой по ряду существенных вопросов, образуя т. о. различные лагери. Часть ученых, ближе примыкающая к самому Ру, стояла на точке зрения, носцвшей преформистский характер. Само развитие регенерата вызывается, по их мнению, раздражением, наносимым ампутацией. Направление Р. определяется в основном под воздействием резервных наследственных зачатков, которые т. о. представляют свойства будущего органа и, попадая при дальнейшем размножении клеток в различные участки регенерата, побуждают их к соответствующему развитию. Большинство из этих исследователей держалось одновременно той точки зрения, что каждая ткань регенерирующего органа образуется за счет подобной же ткани остатка органа, причем их развитие идет до известной степени независимо друг от друга (теория P. «Teil fur Teil»). Преформистекая, каузально-аналитическая теория Р. должна быть решительным образом отвергнута. Она исключает представление о действительном процессе новообразования, трактуя явление как осуществление уже ранее существовавшего. Преформистские представления исходят из предположения, что мы имеем в скрытом виде в лице наследственных зачатков предобразованную структуру будущего органа. Все это предположение носит крайне искусственный характер и стоит в противоречии с современными данными. Также рядом наблюдений было опровергнуто положение о независимом друг от друга развитии отдельных тканей регенерата за счет соответственных тканей культи. Наряду с указанным представлением возникает и другое, обоснование которого принадлежит Дришу (Driesch) и находится в резком противоречии с первым представлением. Дриш принимает, что регенерат не преформирован в регенерирующих частях, иначе пришлось бы предположить наличие в каждой части бесчисленных механизмов, соответствующих различным возможностям развития. Этот вывод основывается на том, что при самых различных уровнях ампутации возникает нормальный орган, следовательно один и тот же участок регенерата может развить в одном случае одно, в другом—другое образование. Дриш считает поэтому, что регенерат является однородным в смысле регенерационной способности отдельных его участков и лишен какой-либо структуры, предопределяющей будущее развитие. Различия между частями будущего органа обусловливаются не различиями частей регенерата, а неодинаковостью положения их в целом (регенерате). Отсюда известное положение Дриша, что судьба части зависит от ее положения в целом. Характер или сущность рассматриваемых различий обусловливается однако не положением в целом, а некиим нематериальным фактором, называемым Дришем энтелехией. Стремления энтелехии направлены на то, чтобы регенерат развивался в нужном для организма направлении. К признанию нематериальности фактора, обусловливающего направление Р., Дриш приходит путем исключения других возможных по его мнению объяснений, которые сводятся к грубо механистическим представлениям. Так. обр., по Дришу, картина регенерационного процесса рисуется в таком виде. Моментом, вызывающим Р., является неопределимое ближе нарушение организма, получающееся в результате ампутации и побуждающее организм к исправлению недостатка. Направление Р. обусловливается энтелехией, действующей целесообразно, и зависит поэтому от конечной цели Р., т. е. формы того органа, который должен образоваться. ■ Несомненная идеалистичность концепций Дриша не мешает ему оставаться механистом. Легко видеть, что метод, применяемый Дришем для объяснения явлений, это тот же каузально-аналитический метод Ру, но на этот раз служащий для обоснования виталистических концепций. Источник развития и у Дриша внешен по отношению к развивающемуся объекту, и развитие анализируется лишь в его формальной обусловленности. В результате такого анализа получается чисто формальное положение о зависимости различий от положения части. Сущность процесса Дриш думает понять, выделив особый фактор, Влияющий на характер явления,—энтелехию. Если в этой части построений Дриша его нельзя обвинить в недостатке хотя бы и формальной логичности, то этого нельзя сказать про его рассуждения по поводу деятельности энтелехии. Здесь сразу бросается в глаза предвзятость и надуманность теории Дриша. Разбив грубо механистическое воззрение и считая, что тем самым исключается всякое материалистическое понимание процесса, Дриш пытается объяснить явление Р. посредством введения нематериального начала. Такая позиция означает однако по существу лишь видимость объяснения, а на самом деле является отказом от последнего; место действительного изучения занимает деятельность воображения.—Уже очень скоро рядом исследований была показана непригодность теории Дриша для объяснения Р. и прямое противоречие ее с наблюдаемыми фактами. Было показано, что регенерационный процесс происходит независимо от того, является ли он целесообразным. Пересаженные органы регенерируют на необычном для них месте, давая там образования, нарушающие гармоничность организма, которая не может т. о. считаться той целью, к которой направлен регенерационный процесс. Вызывание регенерационного процесса на необычном месте путем приведения нерва показывает, что совсем не отсутствие органа является движущим моментом Р. и направление последней стоит в связи не с целесообразным, нематериальным началом, а с вполне материальными свойствами регенерирующей области. Кроме того так как образующийся орган никогда не бывает вполне подобен ранее существовавшему, а иногда и совсем непохож на него до стремление к «восстановлению утраченного» и вовсе может быть оспариваемо. Неудовлетворительность виталистических построений Дриша побудила исследователей искать иного разрешения регенерационной проблемы. В то же время и старое преформист-ское учение было в достаточной мере скомпрометировано. Этим объясняются попытки ио-г строения теорий Р., которые шли бы в ином направлении и были бы лишены недостатков старых. Наиболее разработанные в этом отношении теории принадлежат Гиено и Вейсу и относятся к 20-м годам 20 в. У эпигенетиков эти исследователи заимствуют представление об однородности в смысле потенций регенерационного материала, в то же время они считают, что развитие бластемы определяется тканями, расположенными непосредственно позади регенерата. Таким образом направление развития по мнению этих авторов вносится внешним по отношению к регенерату фактором, с другой стороны, таким фактором оказывается остаток ампутированного органа, т. е. вполне конкретный объект исследования, а не мистический потусторонний фактор, как это имеет место у Дриша. Возможность такого построения достигается тем, что противопоставляются друг другу две различные части регенерата: новообразованные ткани и старые, позади них лежащие. Первые объявляются на основании опытов пересадок лишенными до известного времени специфичности. Наоборот, последняя свойственна старым тканям. Вывод отсюда делается такой, что развитие новообразованных тканей совершается под влиянием старых; первые не обладают самостоятельным, заложенным в них направлением регенерации, оно индуцируется в них позади лежащими тканями, сообщающими бластеме свойственную им структуру. Это основное исходное положение получает то или иное развитие и оттенки в зависимости от того, к какому воззрению примыкает автор. Гиено, более близкий к преформизму, противопоставляет старой эпигенетической точке зрения о зависимости направления Р. от организма как целого идею о том, что организм представляет собой мозаику автономных областей, из которых каждая способна образовать лишь специфический, свойственный ей орган. Такие обособленные части организма Гиено называет «регенерационными территориями». Принимая, что специфичность развития сообщается регенерату позади лежащими тканями, Гиено пытается продолжить анализ и выяснить, какая именно часть этих тканей может считаться ответственной за направление Р. Так как ни одна из использованных в эксперименте тканей (нервы, мускулатура, скелет, кожа) не оказывается специфическим условием Р., то Гиено приходит к выводу, что или приходится приписать это свойство по методу исключения соединительной ткани или связать его с территорией как целым. Какое-либо из этих утверждений было бы с его точки зрения пока преждевременным. Иначе формулирует свои воззрения Вейс, более склоняющийся к эпигенетическим концепциям. Он также принимает, что новообразованные ткани не содержат никакой тенденции к развитию того или иного органа, они «нуллипотентны», неорганизованы. Всякая же организация, по Вейсу, может возникать лишь под влиянием уже организованного материала. Последним являются лежащие позади регенерата части. Влияние организованного материала на неорганизованный происходит не таким образом, что части его влияют независимо друг от друга — организованный материал влияет как целое, он несет «поле». Что такое представляет из себя регенерацион-ное поле по существу, Вейс не разъясняет; он указывает только на нек-рые чисто формальные свойства его, напр. возможность слияния двух «полей» в одно и т. д. Каждая область организма обладает своим специфичным «полем», так. образом организм и по Вейсу представляет собой мозаику «полей». Однако эта мозаика есть результат эмбрионального развития, результат разделения однородного некогда зародыша на независимые части или разделение общего «поля» зародыша на несколько «полей». То разрешение регенерационной проблемы, к-рое дается Гиено и Вейсом, никак не может считаться удовлетворительным. Их ошибка заключается опять-таки в механистичности анализа, в применении каузально-аналитического метода. Направление Р. исследуется ими не в связи с движущими силами регенерациоы-ного процесса, а независимо от них, изучается лишь его формальная обусловленность. Только формальный анализ позволяет делать из того положения, что регенерат до известного стадия неспецифичен, вывод о привнесении направления Р. извне, под влиянием позади лежащих тканей. Это достигается путем искусственного противопоставления частей регенерирую- щего участка, выставления их как внешних друг другу. — Легко показать, что разбираемые теории не разрешают противоречия эпигенетической и преформистской точек зрения. Представление об источнике развития как о части организма, внешней по отношению к рассматриваемому объекту, прямо не дискредитируется лишь, пока мы имеем дело с явлениями Р. Но если, логически продолжая ход рассуждений авторов, поставить вопрос о том, чем определяется развитие в исходном моменте онтогенеза, когда налицо недиференцирован-ное еще яйцо, то мы должны неизбежно или признать наличие какого-то внешнего по отношению к нему фактора или возвратиться к неразрешимым противоречиям прежней преформистской точки зрения. Затруднения, встающие перед разбираемой теорией, естественно сказываются в том, что мы не получаем все же объяснения регенерационному процессу. Гиено вовсе отказывается судить о сущности действия территории, «поле» же Вейса, несмотря на все стремления автора лишить его мистического характера, остается все же не более четким понятием, чем энтелехия Дриша, и несомненно указывает на виталистические тенденции Вейса. Упомянутые до сих пор теории характерны чисто мор фол. подходом к изучаемому объекту. Противоположность этой точке зрения представляет теория физиол. градиентов Чайлда. Чайлд ставит во главу угла своей теории различия в физиол. свойствах разных областей организма. Последние могут быть выявлены различными способами: изучением потребления кислорода, чувствительности по отношению к различным реагентам и т. д. Получающимся при этом количественным различиям Чайлд приписывает решающее значение в смысле влияния на развитие. Степень физиол. активности обусловливает собой появление того или иного образования. Чайлд т. о. заменяет односторонность морфол. точки зрения не менее односторонней физиологической, чисто количественной точкой зрения. Такое разрешение вопроса конечно также неудовлетворительно. Поскольку при Р. дело идет об образовании качественно различных органов, чисто количественное воззрение осуждено ' на бесплодность. И действительно, связь между наличием того или иного градиента и возникновением определенного органа остается у Чайлда неясной. Далее, различия в физиол. активности различных участков имеют, по Чайлду, своим источником определенную область организма, от к-рой исходит необходимое влияние, имеющее энергетический характер. Возникновение же такой «доминирующей» области является результатом реакции протоплазмы на внешний по отношению к ней фактор. Рассматриваемое представление по существу не отвечает на неизбежно встающий вопрос, почему реакция носит именно данный характер. Теория Чайлда носит ту же печать механистичности и формального подхода к явлению, как и ранее разобранные , и поэтому не может дать правильного и непротиворечивого представления о процессе. Таким образом все рассмотренные нами теории Р. не могут быть признаны отвечающими действительности., Они не способны выявить движущие силы явления, моменты, его определяющие, давая неправильное представление о процессе. Вследствие того что исследователи Р. руководствовались ошибочным методом, добытые •18 ими результаты приходится истолковывать совсем иначе, чем они это делают. Приходится отрицать определяющую роль различных факторов," выделенных в результате изучения.Р., и признать эти факторы лишь условиями процесса. На этом представлении нельзя однако ограничиться; т. к. выделение этих условий в большинстве работ протекало с неправильной точки зрения, то выводы авторов могут быть оспариваемы в ряде моментов. С другой стороны, ясно, что нельзя успокоиться на позиции кондиционализма и надо выявить те определяющие отношения, к-рые лежат в основе регене-рационного процесса. Отсюда вытекает необходимость разработки диалектико-материалис-тической теории Р., к-рая одна только может дать глубокое познание явления. В наст, время мы не имеем еще такой теории, однако можно указать, что ее построение предполагает рассмотрение процесса в его самодвижении, не формальный анализ, а вскрытие реальных движущих сил процесса. Л. Лиознер. Регенерация у человека, так же как вообще у всех живых существ, бывает двух типов. А. Нормологическая, или фи зи о логическая Р. имеет место в повседневной нормальной жизни человека и проявляется в непрерывно совершающемся возмещении отживающих тканевых элементов новообразованными клетками. Она наблюдается в той или иной степени во всех тканях, в частности в костном мозгу постоянно идут регенеративные размножения и созревание эритроцитов, возмещающих отмирающие красные кровяные тельца; в покровном эпителии, в к-ром имеет место беспрерывное отъединение ороговевающих клеток, все время совершается возмещение их размножающимися клетками глубоких слоев эпителиального покрова.—Б. Патологическая Р. происходит в результате пат. гибели тканевых элементов. Процесс Р. в последнего рода случаях, собственно говоря, не является пат. процессом; пат. Р. отличается от нормологической Р. не по своему существу, а по своему масштабу и другим особенностям, связанным с характером предшествовавшей убыли тканевых элементов. Так как гибель тканевых элементов в результате различных пат. факторов представляет собой нечто, весьма отличающееся от физиол. отживания клеток как в количественном, так и в качественном отношениях, то отсюда и пат. Р. количественно и качественно ©тличается от нормологической Р. Проявления пат. Р. чаще всего связаны с воспалительным процессом и от последнего они неотделимы резкой границей; строго отграничить, чтб относится к воспалению и что к Р., часто невозможно; в частности проли-феративный фактор в воспалительной реакции весьма трудно отделить от регенеративного размножения клеток. Так или иначе всякое воспаление подразумевает последующую Р., хотя Р., как указывалось, может быть и не связана с воспалением. Ход процесса Р. бывает разным в зависимости от характера повреждения и способа гибели тканевых элементов. Если имело место действие фактора, вызвавшего наряду с повреждением воспалительную реакцию ткани, то обычно проявления Р. начинаются лишь после того, как острый период воспаления, сопровождающийся значительным нарушением жизнедеятельности ткани, стихает. Если в связи с повреждением или в результате развив- шегося воспалительного процесса произошло омертвение ткани, то Р. предшествует или с ней сочетаются про'цессы рассасывания мертвого материала; последние нередко протекают при участии воспалительной реакции. В противоположность этому, если гибель клеток является следствием дегенеративных и атро-фических изменений их, то Р. идет одновременно с этими некробиотическими процессами и не сопровождается воспалением; в частности в печени, в почках наряду с дегенерацией части паренхиматозных элементов можно видеть явления регенеративного размножения лучше сохранившихся клеток; при атрофии одной доли печени от давления, напр. эхинококком, в другой доле идет размножение клеток, нередко полностью покрывающее происходящую убыль печоночной ткани. В основе- Р. лежит размножение клеток, соответствующее нормальному делению их; при этом главное значение имеет непрямое, кариокинетическое (митотическое) деление клеток, тогда как прямое, амитотическое деление наблюдается редко. Кроме картин нормального кариокинеза при пат. Р. могут иметь место пат. формы мйтотического деления в виде абортивных, асимметрических, мультиполярных митозов и пр. (см. Кариокинез). В результате регенеративного размножения клеток образуются юные, незрелые клеточные элементы, которые в дальнейшем созревают, диференциру-ются, достигая той степени зрелости, которая свойственна нормальным клеткам данного вида. Если процесс Р. касается отдельных клеток, то морфологически он выражается в появлении среди ткани отдельных молодых клеточных форм. Если же дело идет о возрождении более или менее обширной тканевой территории, то в результате регенеративного размножения клеток происходит образование незрелой, индифе-рентной ткани зародышевого типа; эта ткань, состоящая в первое время лишь из юных клеток и сосудов, в дальнейшем диференцируется, созревает. Период незрелого состояния регенерирующейся ткани в зависимости от темпа процесса и от различных внешних условий может иметь разную продолжительность. В некоторых случаях весь процесс образования новой ткани идет постепенно, исподволь, причем новые тканевые элементы образуются и созревают не одновременно; при таких условиях, как это напр. бывает при разрастаниях межуточной ткани паренхиматозных органов (печень, почки, мышца сердца) в зависимости от атрофии паренхимы, период незрелого состояния ткани морфологически неопределим. Наоборот, в других случаях, именно, когда ткань данного района подвергается энергичному регенеративному разрастанию, образуется морфологически очевидная незрелая ткань, в дальнейшем созревающая в тот или иной период времени; наиболее демонстративным в этом смысле является разрастание грануляционной ткани. В большинстве регенеративных процессов осуществляется правило сохранения специфической производительности.тканей, т. е. то обстоятельство, что размножающиеся при Р. клетки образуют ту ткань, из которой это размножение исходит: размножение эпителия дает эпителиальную ткань, размножение соединительнотканных элементов образует соединительную ткань. Однако на основании данных о Р. у низших поз- воночных, а по отношению к человеку — данных, касающихся пат. Р., воспалительных разрастаний и опухолей, приходится допустить исключения из этого правила в виде возможности образования в нек-рых случаях из размножающегося и так сказать эмбрионализи-рующегося эпителия тканей мезенхимального характера (соединительной ткани, мышц, сосудов), а из соединительной ткани—развитие мышечных элементов, сосудов, элементов крови. Кроме того при регенерации в определенных тканевых группах (эпителий, соединительнотканные образования) может происходить изменение вида ткани, т. е. то, что именуется метаплазией (см.). Условно принято различать Р. полную и неполную. Полной Р., или реституцией' (restitut-io ad integrum) называют такое возрождение тканей, при к-ром на месте погибшей ткани образуется новая ткань, соответствующая той, к-рая была утрачена, напр. восстановление мышечной ткани при нарушении целости мышцы, восстановление эпителиального покрова при заживлении раны кожи. К неполной Р., или субституции относятся те случаи,, когда дефект не заполняется тканью, подобной бывшей здесь раньше, а замещается разрастанием соединительной ткани, к-рая постепенно превращается в рубцовую ткань; в связи с этим неполная Р. обозначается еще как заживление посредством рубцевания. Очень нередко бывает так, что имеются признаки Р. специфических элементов данной ткани (напр. в поврежденной мышце образование из мышечных волокон «мышечных почек»), однако Р. не идет до конца и дефект замещается преимущественно соединительной тканью. Неполная Р. имеет место при б. или м.. значительных потерях вещества ткани, а также в тех случаях, когда или вследствие особенностей организации поврежденной ткани (см. ниже) или в связи с наличностью тех или иных неблагоприятных условий размножения специфических элементов данной ткани не происходит вовсе или же оно идет слишком медленно; при таких условиях разрастание соединительной ткани получает преобладание. Нужно отметить, что в действительности полная Р. в смысле восстановления ткани, ничем не отличающейся от прежней, нормальной ткани данного места, никогда не наблюдается. Новообразованная ткань, соответствующая в морфол. и фнкц. смысле прежней ткани, все же всегда в той или иной степени отличается от нее. Эти отличия иногда бывают небольшими (недоразвитие . отдельных элементов, нек-рая неправильность тканевой архитектуры); в других случаях они являются более существенными; например образование той же ткани, но упрощенного типа (т. н. гипотипия) или развитие ткани в меньшем объеме. Сюда же относятся случаи суперрегенерации, проявляющиеся у низших животных в образовании лишних органов, конечностей (см. выше), а у человека в так наз. перепроизводстве тканей; последнее заключается в том, что регенеративное разрастание ткани идет дальше границ дефекта и дает избыток ткани. Это наблюдается очень часто, напр. при повреждениях костей, когда избыточно новообразованная костная ткань выступает в виде утолщений, выростов, иногда весьма значительных; при Р. в эпителиальных покровах и железистых органах, когда размножающийся, эпителий образует весьма значительные разрастания, приближающиеся к проявлениям опухолевого роста, напр. атипические разрастания эпителия при Р. язв и ран кожи и слизистых оболочек, регенеративные аденомы в печени и почках при заболеваниях этих органов, сопровождающихся гибелью части их паренхимы. В большинстве случаев такая избыточно разросшаяся ткань бывает лишена фнкц. значения; иногда (в костях) она . в дальнейшем подвергается убыли путем рассасывания. Условия Р. у человека очень разнообразны и сложны. Среди них большое значение имеют те весьма многочисленные факторы с которыми связаны реактивные способности организма вообще; сюда относятся наследственно-конституциональные особенности организма, возраст, состояние крови и кровообращения, состояние питания и обмена веществ, функция эндокринной и вегетативной систем, а также и условия жизни и труда индивидуума. В зависимости от установок этих факторов Р. может итти тем или иным темпом, с той или иной степенью совершенства; у разных индивидуумов при повреждении одинакового типа Р. ткани может протекать нормергически, гиперергически, анергически или же вовсе отсутствовать. Важное значение для Р. имеют и местные условия со стороны той области, где происходит Р.: состояние в ней кровообращения, лимфообращения; отсутствие или наличие воспаления, особенно нагноения. Само собой понятно, что образование новых клеток может происходить лишь при достаточной! подвозе кровью питательного материала; далее размножение и созревание клеток не может происходить в тканях, находящихся в состоянии резкого воспаления.—Очень существенное значение для Р. имеет характер регенерирующейся ткани в смысле степени ее организации и специфической диференци-ровки, а также других особенностей строения и существования ткани. Чем выше развитие ткани, чем сложнее ее организация и диферен-цировка, чем специальнее ее функция, тем в меньшей степени ткань способна к Р.; и, наоборот, чем менее сложно построена и диферен-цирована ткань, тем в большей степени ей свойственны регенеративные проявления. Это правило обратной пропорциональности между способностью тканей к Р. и степенью их организации не является однако абсолютным; кроме степени диференцировки всегда имеют значение и другие биол. и структурные особен-v ности ткани; напр. клетки хряща гораздо в меньшей степени способны к Р., чем более сложно организованные клетки эпителия. В общем все же можно отметить, что напр. мало диференцированные клетки соединительной ткани, клетки покровного эпителия обладают большой способностью к Р., тогда как возможность регенеративного размножения таких высоко диференцированных элементов, как нервные клетки головного и спинного мозга, как мышечные волокна сердца, до сих Нор еще не доказана и сомнительна. На середине стоят клетки секреторного эпителия железистых органов и волокна произвольной мускулатуры, которым свойственна Р., но далеко не такая совершенная, как соединительной ткани и » покровного эпителия. То обстоятельство, что регенеративное размножение в большей степени свойственно менее зрелым и развитым клеткам, проявляется еще в том, что во вся- . кой ткани регенерация исходит из тех ее зон, в к-рых сохраняются менее зрелые элементы (в покровном э'пителии из базального или герминативного слоя, в железах — из .назальных частей выводных протоков, в кости—из эндоста и периоста); эти зоны принято называть проли-ферационными центрами или центрами роста. Регенерация отдельных тканей. Р. крови, напр. после кровопотерь, происходит таким образом, что сначала путем диффузии и осмоса через сосудистую стенку восстанавливается плазма крови, после чего в крови появляются новые, красные и белые кровяные тельца, к-рые возрождаются в костном мозгу и в лимфаденоидной ткани (см. Кроветворение).-—-Р. кровеносных сосудов имеет важное значение потому, что она сопровождает Р. всякой ткани. Существуют два типа образования новых сосудов.—А. Чаще всего имеет место почкование старых сосудов, к-рое заключается в том, что в стенке мелкого сосуда происходит набухание клетки эндотелия и кариокинетическое деление ее ядра; образуется выбухающая кнаружи как бы почка (образование т. н. ангиобласта), к-рая в дальнейшем при продолжающемся делении ядер эндотелия вытягивается в длинный тяж; в последнем в направлении от старого сосуда к периферии появляется просвет, благодаря чему бывший сначала массивным тяж превращается в трубку, начинающую пропускать кровь. Образующиеся таким образом новые сосудистые веточки соединяются друг с другом, что дает образование сосудистых петель.—Б-. Второй тип новообразования сосудов называется аутогенным развитием сосудов. В основе его лежит образование сосудов непосредственно в ткани без связи с прежними сосудами; непосредственно среди клеток появляются щели, в к-рые открываются капиляры и изливается кровь, причем прилегающие клетки получают все признаки эндотелиальных элементов. Такой способ, сходный с эмбриональным развитием сосудов, может наблюдаться в грануляционной ткани, в опухолях и повидимому в организующихся тромбах. В зависимости от условий кровообращения новообразованные сосуды, имевшие сначала характер капиляров, в дальнейшем могут приобретать характер артерий и вен; образование прочих элементов сосудистой стенки, в частности гладких мышечных волокон, в таких случаях идет за счет размножения и диференцировки эндотелия. Образование новой соединительной ткани имеет место в качестве регенеративного проявления при повреждениях самой Соединительной ткани и кроме того как выражение неполной Р. (см. выше) самых разнообразных других тканей (мышечной, нервной и пр.). Кроме того новообразование соединительной ткани наблюдается при весьма разнообразных пат. процессах: при т.н. продуктивных воспалениях, при исчезании паренхиматозных элементов в органах вследствие их атрофии, дегенерации и некроза, при заживлении ран, при процессах организации (см.) и инкапсуляции (см.). При всех этих условиях сначала происходит образование юной, незрелой грануляционной ткани (см.), подвергающейся созреванию до степени зрелой соединительной ткани. —Р. жировой ткани происходит из ядросодержащих остатков протоплазмы жировых клеток или же путем превращения в жировые клетки обычных клеток соединитель- ной ткани. В том и другом случае сначала образуются округлые клетки—липобласты, протоплазма к-рых выполнена массой мелких жировых капелек; в дальнейшем эти капельки сливаются в одну крупную каплю, отодвигающую ядро к периферии клетки. Р. костной тк.ани при повреждениях кости имеет в основе размножение остеобластов эндоста и камбиального слоя периоста, к-рые вместе с новообразованными сосудами образуют остеобластическуюгрануляционную ткань. При костных переломах (см.) эта остеобластическая ткань формирует т. н. провизорную (предварительную) костную мозоль. В дальнейшем между остеобластами появляется плотное,- однородное вещество, благодаря чему новообразованная ткань приобретает свойство остеоид-ной ткани; последняя, петрифицируясь, превращается в костную ткань. При переломах это совпадает с образованием дефинитивной (окончательной) костной мозоли. При фнкц. нагрузке устанавливается определенная архитектура новообразованной костной ткани, что сопровождается рассасыванием лишних частей и образованием новых (перестройка кости).— Хрящевая ткань способна к Р. в сравнительно слабой степени, причем хрящевые клетки в регенеративных проявлениях участия не принимают. При небольших повреждениях хряща происходит размножение клеток глубокого слоя надхрящницы, называемых хонд-робластами; вместе с новообразованными сосудами эти клетки образуют хондробластйчес-кую грануляционную ткань. Между клетками последней вырабатывается основное вещество хряща; часть клеток «атрофируется, исчезает, другая часть превращается в хрящевые клетки. Крупные дефекты хряща заживают рубцеванием.—Р. мышечной ткани—см. Мышцы. Эпителиальная ткань, особенно покровный эпителий кожи, слизистых оболочек, серозных покровов, в высокой степени способна к Р. При дефектах в многослойном плоском эпителии кожи и слизистых оболочек образуется новая эпителиальная ткань, являющаяся продуктом кариокинетического деления клеток зародышевого слоя сохранившегося эпителия. Образующиеся юные эпителиальные клеточки надвигаются на дефект и покрывают его сначала одним слоем низких клеток; в дальнейшем при продолжающемся размножении этих клеток формируется многослойный покров, в к-ром идет созревайие и диференцировка клеток, соответствующая структуре обычного многослойного плоского эпителия. На слизистых оболочках, покрытых цилиндрическим эпителием, дефекты замещаются, надвигающимися эпителиальными клеточками, являющимися продуктами размножения клеток сохранившихся желез (в кишечнике—Либеркюнрвых, в матке—маточных желез); здесь точно так же дефект сначала покрывается низкими, незрелыми клетками, к-рые в дальнейшем созревают, делаются высокими, цилиндрическими. При Р. слизистой оболочки матки и кишечника из такого эпителиального покрова при продолжающемся размножении его клеток образуются трубчатые железы. Плоский эпителиальный покров серозных оболочек (брюшины, плевры, перикарда) восстанавливается при посредстве кариокинетического деления сохранившихся клеток; при этом в первое время новообразованные клетки имеют более крупные размеры и кубическую форму, а потом уплощаются. ■Й57 По отношению к Р. железистых органов надо отличать, с одной стороны, гибель и возрождение лишь железистого эпителия при сохранении основной структуры органа, а с другой стороны—повреждение с последующей Р. всей ткани органа в целом. Р. эпителиальной паренхимы железистых органов после частичной гибели ее вследствие некроза и перерождений происходит весьма совершенно. При различных перерождениях и некрозе, напр. эпителия печени, почек, сохранившиеся клетки подвергаются кариокинетическому (реже прямому) делению, благодаря чему и происходит замещение утраченных элементов равноценными железистыми клетками. Возрождение частей железистых органов в целом более •сложно и в общем бывает очень редко совершенным. В нек-рых железах, напр. щитовидной железе и в слезных железах, иногда наблюдают образование отпрысков от сохранившейся железистой ткани и образование новых железистых ячеек. В других органах возрождение бывает гораздо более слабым; часто над ним преобладают процессы гипертрофии и гиперплазии сохранившихся эпителиальных элементов. В. частности в печени при гибели ее ткани происходит размножение и одновременно увеличение объема печоночных клеток только в пределах сохранившихся долек; на разрезе такой печени невооруженным глазом в соответствующих местах часто бывает заметен более крупный рисунок строения долек. В общем такие процессы размножения и увеличения объема клеток в сохранившейся печо-ночной ткани могут достигать очень большой степени; есть наблюдения, указывающие, что при постепенном изъятии 2/3 частей печени сохранившаяся треть ее может дать увеличение объема, покрывающее вышеуказанную убыль. В противоположность этому образования" новой печоночной ткани в целом, т. е. новых долек с их системой капиляров и пр., никогда не наблюдается. Очень часто имеет место новообразование желчных протоков, дающих многочисленные новые веточки; на концах последних клетки часто подвергаются увеличению в объеме и начинают напоминать печоночные клетки, но дальше этого развитие их не идет. В почках при гибели их ткани, напр. при образовании инфаркта, новая почечная ткань не образуется вовсе; лишь иногда наблюдается образование небольших отпрысков от канальцев. Вместе ■с тем может происходить увеличение объема клубочков и канальцев в сохранившихся отделах почки. При Р. эпителиальной ткани нередко происходит значительная перестройка ее, т. е. изменение формы и взаимоотношений структурных частей. Иногда имеет место метаплазия; часто встречгется перепроизводство ткани в виде атипических разрастаний эпителия (см. выше). В нервной ткани Р. в очень разной степени касается собственно нервных элементов и невроглии. Возрождения погибших нервных клеток в сформированной центральной нервной системе человека повидимому не происходит вовсе; лишь изредка описывались- не вполне убедительные картины как бы начинающегося деления ядер этих клеток. Ганглиозные клетки симпат. нервной системы в молодом организме могут размножаться, однако это имеет место весьма редко. Все потери вещества в центральной нервной системе заживают посредством заполнения дефекта разрастающейся тканью нев- роглии, которая в высокой степени способна к регенеративным проявлениям, особенно т. н. мезоглия. Кроме того крупные дефекты в мозговой ткани могут выполняться соединительной тканью, разрастающейся из мозговых оболочек или из окружности кровеносных сосудов. Р. периферических нервов—см. Нервные волокна, регенерация нервных волокон. а. Абрикосов. Лит.: Астрахан В., Материалы к изучению закономерностей в процессе регенерации, Москва, 1929; Давыдов К., Реституция у немертин, Труды Особого зооп. каб. и Севастопольской биол. станции, Академия наук, серия 2, № 1, 1915; Леб Ж., Организм как целое, Москва—Ленинград, 1920; Korschelt E., Regeneration und Transplantation, Band I, Berlin, 1927; Morgan Т., Regeneration, New York, 1901; Scha-xel J., Untersuchungentiber die Formbildung der Tiere, Band I — Auff assungen und Erscheinungen der Regeneration, Arb. aus dem Gebiete der experiment. Biologie, Heft 1, 1921. Смотрите также:
Смотрите также:
- REGIO SUBTHALAMICA (подбугровая область; CHH.:regio hypothalamica,hypothalamus), нижняя часть межуточного мозга (diencepha-lon), расположенная книзу от зрительного бугра и образующая нижнюю часть стенки III желудочка и его дно. Развивается из переднего первичного мозгового пузыря, ...
- РЕГИСТРАЦИЯ болезней представляет начальный акт в деле статистического исследования заболеваемости населения. Задачей ее является собирание первичного материала. Р. заключается в учете каждого отдельного случая проявления исследуемого явления и описании его по ...
- РЕГРЕССИЯ, количественные изменения одного явления, наступающие в связи с изменениями другого явления, находящегося с первым в корреляционной связи. Так, последовательные изменения веса в отдельных частях какого-либо однородного по основным признакам ...
- РЕДУЦИРОВАННОЕ КРОВООБРАЩЕНИЕ, понятие, введенное Оппелем в 1911 году для обозначения такого состояния, когда конечность живет на счет колятерального кровообращения (как артериального, так и венозного) в тех случаях, когда к вынужденной перевязке ...
- РЕДУЦИРОВАННЫЙ ГЛАЗ, условная оптическая система, обладающая теми же свойствами в смысле преломления лучей, как и настоящий глаз, но значительно более удобная для всякого рода расчетов. Как известно, всякая система сферических оптических ...