ПРОПРИОЦЕПТИВНЫЕ ЭЛЕМЕНТЫ
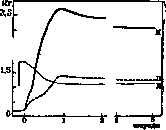
Эти рефлексы играют немалую роль при поддерживании и перемещении грузов; при держании груза одновременно охвачено возбуждением только такое количество миофибрил, напряжения которого достаточно для уравновешивания груза. При поднимании же груза сокращаются кроме того и невозбужденные этим механизмом миофибрилы. Миостатические ре^ флексы наблюдаются при длительном растяжении мышц. Гофман (Hoffmann) выделил в особую группу «собственных рефлексов» (Eigenre-flexe) те, которые обусловлены мгновенным. растягиванием мышцы и состоят в одиночном вздрагивании мышцы в ответ на ее растяжение, Гофман относит к этой группе явлений все сухожильные рефлексы, считая, что удар по сухожилию действует только как растягивание мышцы. При этом интенсивность сухожильных, рефлексов различна—она выше всего в разгибателях ниж- 3-° них конечностей 2,5. и на сухожили- 2,о. ях мышц пальцев и пронато-рах руки (в этих 10 же мышцах лег- o,s че всего наступает контрактура при описанной Вернике форме гемиплегии). На интенсивность сухсжильн. рефлексов влияют самые разнообразные факторы, как нормальные
(утомление, гипервентиляция), так и патологические (например tabes dorsalis). Диагностическое значение сухожильных рефлексов—см. Рефлексы. По Гофману «собственные рефлексы» играют роль в процессах нормальной иннервации мускулатуры, выступая как «регулятор» интенсивности мышечных сокращений: они усиливают сокращение мышц при держании груза и умеряют интенсивность сокращения при разгрузке мышцы. Показано, что ритм произвольной иннервации (ритм токов действия мышц) нарушается при выпадении «собственных рефлексов». Делать однако окончательные выводы об их значении преждевременно. Необходимо также отметить, что Фер-стер (Foerster) возражает против представления о сухожильных рефлексах, как о результате только одного растяжения мышцы, и не считает исключенным участие чувствительности: периоста. Он точно так же отделяет сухожильные рефлексы от «рефлексов на растяжение» (Dehnungsreflexe)—сопротивления, рефлектор-но оказываемого мышцей ее не мгновенному растяжению. В последнюю группу явлении входят миостатические рефлексы Шеррингтона. Выключение П. иннервации путем кокаи-низации мышцы нацело выключает «рефлексы на растяжение» (Magnus, LiIjestrand).—Наряду с участием импульсов от П. э. в координации движений эти импульсы играют неменьшую, подчас определяющую роль в мышечном тонусе. Шеррингтоном доказано, что тонус мышц совершенно исчезает после перерезки их чувствительного нерва. Раздражения с кожи и суставов могут усиливать или ослаблять тонус; Рисунок 1. Изменение напряжениями m. quadricipitis при растяжении на 8 мм. Сплошная линия (М) показывает изменение напряжения в нормальной мышце; прерывистая линия (П)—в мышце после перерезки идущего к ней нерва; пунктирная линия (Д)— изменение длины мышцы (изменение длины в пределах 0—8 мм). обеспечить его поддержание при выключении ТЕ. э. они не в состоянии. Дальнейшие исследования Магнуса и его 'школы (de Kleijn, Ra-demaker) подтвердили значение П. э. в поддержании позы, в регуляции мышечного тонуса (см. Тонус). Надо также указать, что про-приоцептивным элементам шейных мышц принадлежит особая роль в регуляции тонуса (см. Магнус-Клейна рефлексы). До сих пор речь шла о нек-рых формах деятельности, обусловленных возбуждением П. э. В живом организме описанные реакции бесспорно находятся в постоянном взаимодействии; напр. при нек-рой средней степени тонического напряжения мышц сухожильные рефлексы в них выражены всего резче; миостати-ческие рефлексы имеют большое значение в поддержании тонуса (но не исчерпывают его, т. к. тонус наблюдается не только в растягиваемых мышцах); в процессе рецяпрокной иннервации, обусловленной возбуждением П. э., меняется «тонус мышц и т. д. Необходимо также помнить, что результат раздражения П. э. может резко меняться при изменении состояния и текущей работы нервных центров. Так напр. раздражение п. peronaei тормозит миостатический рефлекс; реципрокная иннервация антагонистов может нацело меняться при возникновении новых межцентральных отношений [например при доминанте—см. Доминанта(в физиологии)] или просто при изменении типа движения и падающей на мышцы нагрузки. Сколько-нибудь полной картины роли П. э. в координации различных движений дать еще нельзя* как нельзя дать точных данных о характере процессов, происходящих В П. э. и соответствующих нервных центрах. Не приходится однако сомневаться в том, что П. импульсы играют огромную роль в координации позы (сидение, стояние) и движения. Всякая поза поддерживается только при закономерном распределении тонических возбуждений в соответствующих мышечных группах. Разумеется, в нормальной деятельности импульсы, идущие от П. э., действуют в комплексе с импульсами, идущими от рецепторов кожи и высших органов чувств (гл. обр. зрения). Вероятно однако, что в рефлекторной регуляции мышечных напряжений, в координации автоматизированных движений роль П. э. выступает на первое место. Изолированное патологическое поражение только одних П. э. повидимому не встречается, целый же ряд пат. симптомов бесспорно связан с поражением чувствительных элементов центр, нервной системы, воспринимающих раздражение П. э. Наиболее простым случаем является почти полное исчезновение «рефлекса на растяжение» и тонуса после поражения задних корешков спинного мозга (tabes dorsalis). В этом случае также наблюдается атаксия, наблюдаемая однако и при ряде других поражений нервных центров (напр. поражениях мозжечка—Luciani, Goldstein, Foer-ster). Мы должны представить себе дело таким образом, что в пределах одного спинного мозга уже заложены основные механизмы тех рефлекторных изменений в мускулатуре, к-рые обусловлены раздражением П. э. Высшие же отделы мозгового ствола могут разнообразно изменять результат этих раздражений. Полный анализ этих взаимодействий различных отделов центральной нервной системы при нарушении координации движений пока дан быть не может. Несколько более просты зависимо- сти для частного случая П. рефлексов—-рефлексов на растяжения, т. е. вышеупомянутого сокращения мышц при их растяжении, вызываемом напр. пассивным разгибанием конечности; при субъективной оценке наступающее сокращение регистрируется как сопротивление, оказываемое мышцами; при записи токов действия при этом наблюдаются отчетливые электрические явления в мышце. Рефлекс на растяжение (Dehnungsreflex) наблюдается при сохранении только одного спинного мозга с нормальной чувствительной иннервацией. Поражения мозжечка вызывают резкое ослабление или выпадение этого рефлекса. В противоположность этому поражение красных ядер приводит к исключительно резкому усилению рефлексов на растяжение. Сопротивление мышц растягиванию при этом настолько велико, что для сгибания вытянутой конечности требуется значительное усилие (после же достигнутого сгибания конечность «застывает» в приданном ей новом положении). Это крайнее повышение тонуса охватывает гл. обр. мышцы разгибатели. Оно наблюдается во всех случаях, когда красные ядра отделены от нижележащих отделов, среди которых однако обязательно присутствие продолговатого мозга: разрез спинномозговой оси ниже calamus scriptorius не вызывает этого резкого усиления тонуса разгибателей, а всякий разрез выше этого места до уровня красных ядер приводит к вышеописанному явлению. Дело идет о том, что красные ядра посылают какие-то импульсы на нижележащие отделы, препятствующие развитию усиленного тонуса разгибателей («децерзбраци-онной ригидности» Шеррингтона). В клинике описаны случаи опухолей в районе красных ядер, как бы отрезавшие их от нижележащих центров; при этом наблюдалась картина, весьма близкая к той, к-рая найдена при экспериментальной перерезке мозга ниже nuclei rubri; вероятнее всего красные ядра каким-то образом тормозят П. рефлексы, наступающие в ответ на растягивание мышцы. Что же касается коры головного мозга, то ее поражение или поражения пирамидных путей приводят лишь к временному падению интенсивности рефлексов на растяжение. Подробнее же роль кортикальной иннервации в протекании и регуляции П. рефлексов остается, как сказано, неизвестной. На примере «рефлексов на растяжение» видно многообразие иннервационных факторов, регулирующих даже сравнительно простую форму П. рефлексов. Полная же картина регуляции П. иннервации не может быть дана, хотя мы имеем все основания рассматривать последнюю как играющую основную роль во всей координации дви- ^g_ гательных актов. Их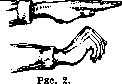
резкое расстройство при поражении П. иннервации хорошо демонстрируется приведенным рисунком (рис. 2). Больной с полным поражением П. чувствительности правой руки- не может при закрытых глазах придать обеим рукам одинаковый уровень и положение.
Наряду с значением П. э. как органов, ре-флекторно регулирующих мышечную деятельность, надо указать на их значение как органа восприятия чувства тяжести и положения тела. Действующие на мышцы (через кожу и костно- связочный аппарат) грузы стремятся их растянуть, что вызывает' вышеописанные процессы в П. э. При этом точность оценки груза при динамическом сокращении мышц выше, чем при их статическом напряжении для . удержания груза. Необходимо признать, что П. импульсы доходят до коры мозга и воспринимаются сознанием. При обычных движениях П. импульсы повидимому в меньшей степени доходят до коры мозга, так как они направляются в другие нервные механизмы. При фиксировании же внимания (например при оценке груза) на раздражениях от П. э., а также при переходящей за некоторый индивидуальный предел величине груза, раздражение П. э. воспринимается с полной отчетливостью. Вероятно, что некоторые формы мышечного утомления имеют в своей основе изменения в воспринимающих аппаратах головного мозга под влиянием чрезмерно частых (или сильных) П. импульсов. Бесспорно также, что чувствительность корковых отделов (и всей центральной нервной системы) к П. возбуждениям зависит от состояния нервных центров. Лит.: СтарлингЭ., Основы физиологии человека, М.—Л., 1931; ЭдрианЕ., Основы ощущений, М.—Л., 1931; Hoffmann P., Untersuchungen iiber die Eigen-reflexe (Sehnenreflexe) menschlicher Muskeln, В., 1922; Liddell E.u.SherringtonC, Reflexes in response to stretch (myostatic reflexes), Proc. roy. soc, v. XCVI, p. 212—242, 1924—25; они же, Further observations on myostatic reflexes, ibid, p. 267—283; Sherrington C, The integrative action of the nervous system, N. Y., 1906. Г. Конради. Смотрите также:
Смотрите также:
- ПРОПУЛЬСИЯ (от лат. propellere—гнать вперед), симптом, характеризующийся тем, что больной при ходьбе непроизвольно начинает двигаться все быстрее и быстрее вперед, достигая почти скорости бега; если ему приказать остановиться, то он не ...
- ПРОСТАТИН, Prostatin (син. простатокрин, Prostatocrin), препарат, получаемый из предстательной железы быков. В продаже имеется в виде порошка, таблеток (по 0,3—0,5), в ампулах (по 1—2 см3) для подкожных инъекций и в ...
- ПРОСТАТИТ (prostatitis), воспаление, предстательной железы; возникает в результате проникновения в нее микроорганизмов, чаще всего гонококка. Иногда имеет место смешанная инфекция (стафилококки, стрептококки, В. coli и др.). По статистике Социна и Буркгардта ...
- ПРОСТАТОРЕЯ, выделение из уретры во время мочеиспускания или испражнения нормального или измененного секрета предстательной железы без примесей сперматозоидов. Выделение при тех же условиях семени из семенных пузырьков носит название сперматореи ...
- ПРОСТИТУЦИЯ (от лат. prostituere—публично выставлять, ставить на продажу). Проститутка—это женщина, продающаяся мужчинам как объект полового сношения с ними. Половая жизнь женщины, промышляющей продажей своего тела, почти всегда характеризуется беспорядочными сношениями с, ...