ПОЛ

можно констатировать различие в строении клеток разных индивидуумов; нити, обладающие крупными клетками, повидимому соответствуют женским, а нити с более мелкими клетками—мужским особям. При копуляции содержимое мужских клеток перетекает в женские, в результате чего образуются яйцевидные зиготы, каждая из которых окружается плотной оболочкой и дает начало новой нити спирогиры. Однако Гартман (Hartmann) показал, что отдельные экземпляры спирогиры ведут себя одновременно как мужская и как женская особь: при копуляции трех нитей друг с другом (рис. 1) средняя нить В функционирует по отношению к нити А как мужская, а к нити С — как женская. У более высоко организованных растений и животных раздельнополость проявляется вполне отчетливо прежде всего в форме и поведении половых клеток. Наиболее общей характеристикой гамет обоих полов является относительная неподвижность и большая величина яйца по сравнению со спермием, который отличается подвижностью и снабжен различно устроенными органелла-ми движения (реснички, жгуты, мембраны). Появление различно у разных полов устроенных гамет связано с наличием органов многоклеточного организма, являющихся местом образования половых клеток. Половые железы, семенники самцов и яичники самок, называются иногда первичными половыми признаками.
Численное соотношение полов. Наиболее разработанные данные по численному отношению П. относятся к человеку. На 100 родившихся живыми девочек приходится в среднем 105 мальчиков. Вариации для различных стран весьма незначительны. Так, по Бодио (Bodio), для периода 1887—95 гг. на 100 девочек рождается мальчиков (живорожденные): Испания..... Ю8,4 Австрия..... 105,8 Италия ..... 105,8 Голландия ... 105,5 Россия ..... 105,4 Германия .... Юо,2 Для различных животных численное соотношение полов таково: J3 С 1. Копуля-трех нитей спирогиры (из Гартмана). А Рис ция Венгрия . Франция . Швеция . Бельгия . . . 105,0 . . 101,6 . . 104,5 . . 101,5 Англия . . 10.{,6 Животное На 100 самок рождается самцов Автор Рогатый скот . . Голуби ..... Двукрылые . . . 101,0; 98,3* 104,6 115,4; 97,7 104,9; 111,8 105,0 94,7 115,0 105,5 Ь5,8 Корвевен, Дю- зинг Корневен, Дю- зинг Ирвин Корневен, Виль- кенс Кено Дарвин Кено Штандфус Кено * Две рядом стоящие цифры относятся к данным разных авторов. Т. о. численное отношение П. обычно мало уклоняется от равенства. Для получения достоверных результатов необходимо производить подсчет П. у зародышей (если это возмож- но) или у недавно родившихся детенышей. Иначе при определении численного отношения П. в природе можно столкнуться с значительными отклонениями от равенства в силу различных биол. особенностей того или другого вида. Так, у паука Latrodeotes mactans на 100 самок приходится 819 самцов, у рыбы Lophius piscatorius— 385, у рыбы Cottus gobio—183, у черного стрижа—400, у жука Maerodartylus—131; наоборот, у моллюска кальмара (Loligo) на 100 самок приходится 16,6 самцов и у осьминога (Octopus)— 13,3. Такие нарушения равного отношения П.отражают не картину рождения их в таком отношении, а результат или большего процента гибели одного П. или трудность обнаружения одного из П. вследствие его малой заметности (покровительственная окраска) или скрытого образа жизни или же иных биол. условий (для птиц например неодновременность пролета самцов и самок на зимовку или в места гнездования). В большинстве случаев причины уклонения от нормального численного отношения П. в природе могут быть установлены, и можно принять за правило, что при рождении количество самцов и самок одинаково. (Объяснение этого обстоятельства см. ниже—Определение пола.) Половые признаки, У многих животных самец отличается от самки весьма незначительно, а иной раз по внешнему виду различить их невозможно. Однако по строению половых органов у таких мономорфных видов П. нетрудно определить. Помимо различия в анат. и гист. строении половых желез самец мономорфных видов обычно отличается от самки устройством половых проводящих путей и нередко совокупительных приспособлений. Последние обладают особенно сложным строением у насекомых, в частности у бабочек. Кроме органов, играющих непосредственную роль в процессе оплодотворения и обеспечивающих проникновение сперматозоидов в половые проводящие пути самки, известно много приспособлений вне полового аппарата, связанных с актом оплодотворения. Сюда относятся брачные мозоли на большом пальце самца лягутнки, служащие ему для обхватывания и удержания самки во время икрометания. Своеобразные приспособления имеются у нек-рых прямокрылых. У сверчков из рода Oecanthus самец имеет на спине 5 ямок, образованных вдавлением хитинового покрова, в к-рые открываются протоки т. н. ханкокковских желез. В момент спаривания ямки наполнены жидким секретом желез, и самка, сидящая во время копуляции на спине самца, пьет эту жидкость. После копуляции самка открывает прикрепленную к ее половому отверстию сперматофору и съедает ее. Пищевая приманка-в виде секрета ханкокковских желез имеет очевидно значение задержать более долгое время сперматофору у полового отверстия самки, дабы сперма могла перейти в ее половые пути. К числу признаков, отличающих самца от самки у насекомых, относятся приспособления для издавания звуков. Такие приспособления имеются у самцов сверчков, кузнечиков, цикад и др. Им приписывалась роль привлечения самок, но положение это остается недоказанным. У самки тутового шелкопряда (Bombyx mori) и многих других бабочек на конце брюшка имеются железы, вырабатывающие пахучее вещество, несомненно привлекающее самцов, т. к. они собираются вокруг бумажки, на которую выдавлено содержимое пахучих желез, или во- 16& круг кокона с еще невылупившейся самкой. У многих насекомых и позвоночных животных самцы обладают органами, служащими им как оружие при драках во время брачного периода и для защиты от врагов. Сюда относятся рога оленей, клыки кабанов и бабируссы, клыки моржей, бивни слонов и т. п. Описанные здесь признаки постоянно отличают самца от самки; имеется однако много примеров появления отличительных признаков только на время раз- Рис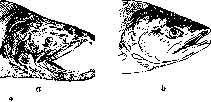
Голова самца кеты в брачном наряде (а); Ъ— внебрачного периода.
множения, т. н. брачный наряд, особенно отчетливо заметный у нек-рых рыб. У самца лосося во время икрометания на нижней челюсти развивается загнутый отросток и вырастают крупные заостренные и изогнутые зубы. У тихоокеанских лососевых рыб. горбуши и кеты, во время нереста (икрометания) серебристая окраска очень сильно темнеет, появляются красноватые пятна и полосы, челюсти сильно удлиняются и снабжаются крупными зубами. Все эти признаки исчезают после брачного периода (рисунок 2). Яркую окраску во время нереста приобретают нек-рые другие рыбы, напр. колюшка (Gasterosteus), гольян (Phoxinus) и др. Появление брачного наряда имеет место и у нек-рых птиц. Здесь его сохранение не ограничивается только периодом размножения, в брачном пере птица остается иногда более полугода. Так, дикий селезень (Anas boschas) зимой и весной несет яркое перо, резко отличающее его от самки, а летом и осенью имеет скромную окраску, напоминающую окраску самки. Б.рачные мозоли самцов лягушек увеличиваются в период икрометания и т. о. должны быть частично отнесены к сезонным половым признакам. Что касается явления постоянного полового диморфизма, то проявления его в мире животных чрезвычайно разнообразны. По существу весь организм самца отличается от самки, но половые отличия особенно отчетливо проявляются в ряде специальных признаков, получивших название вторичных половых признаков (си.). Из анат. особенностей, отличающих самца от самки у постоянно диморфных видов, можно отметить следующие. У червя Bone Ilia viridis самка имеет тело величиной со сливу и хобот, способный вытягиваться, до 1 м длиной, а самец, живущий внутри самки, достигает в длину нескольких миллиметров (рис. 3). У многих бабочек окраска крыльев самца отличается большей яркостью, чем окраска самки; иногда самки вовсе лишены крыльев. У жукоз самец часто значительно крупнее самки и имеет различные хитиновые придатки в виде непарных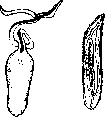
Рисунок 3. Рисунок 4.
Рисунок 3. Bonellia viridis: a — самка (уменып. в 2 раза); Ь—самец (увелич. в несколько раз). Рисунок 4. Нижняя гортань утки (а) и селезня (6). или парных рогов. У многих живородящих рыб есть формы, самец которых обладает яркой расцветкой, отсутствующей у самки, например гупия (Lebistes reticulatus), меченосец (Xiphophorus Helleri) и др. Особенно разнообразно проявляется постоянный половой диморфизм у птиц. Очень отчетливо он выражен в окраске кур, уток, фазанов, павлинов, африкан-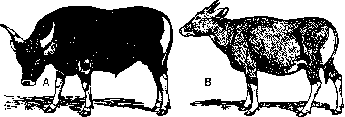

Рисунок 5. Бык (А), корова (В) и кастрированный бык (С) баптенги; бык (D), короьа (F) и вол (Е) серого украинского скота. (Из Завадовского.)
ских страусов, а также у райской птицы, различных видов колибри и т. п.—Одним из замечательных признаков полового диморфизма является устройство голосового аппарата и пенье у многих воробьиных птиц. Нижняя гбр- •6

Ърг \ш0Ь <W
^1 'V*.
tf ---------—?*------------
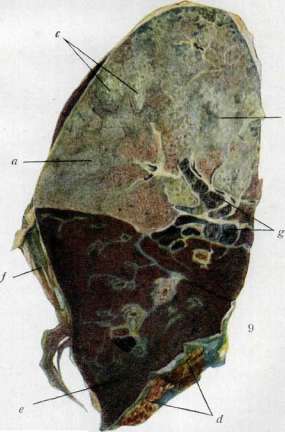
Рисунок I. I lopuNj.ijjiii.tit iit-ryn. Рис, I, КвстрнроиЯнНин нетук■ Pet <f< КжтрнровШшип пстук С lifi'L-L-i.-hi-iuiM^i ему ДООШКОН. Рнс. Ч. Нормальная Kvp)inp. 1*нс, 5. Кшмриронаннап курнпи. Глс. IV Квсгрчронйнлым курица с переевнйнш^м сЕ1 СТГИЯНЮНГОН. Рисунок 7. Фиброзные эксудат f d/ в олыдолач при гглсьмовнн; й — сгеяиг пльнеоли, Рисунок Л. Обилие Фрнллснлероисккх лнпло-баннлл faj и пяевмошнесшж МССудатс; Л— резкая гиперемия сосудов, Рис, 9* Круподоен пвелыоляя (серое опечецелне) с ]!■:.!■ !■■■ ti : i.::.....-: ■ i. i - n пулктад Ь кс скоплений гнойного жсулпта, легко выд анл 1тш,чмого iii пнренмтчи (а)\ d—плевра тичсскнМ эксудат л и сняли с л ни ателектаз нижней доли (c}\f— старик ллеврнтпческне спайки: л,г—антракоз бронхиолы с ик жь'к.]. К ст. Пнсп.ияний, ffa,i. тань (syrinx) у самцов певчих и нек-рых других птиц имеет иное устройство, чем у самки; иногда она снабжена расширениями, играющими роль резонаторов (рис. 4). Поведение самцов в период размножения (ток глухарей и нек-рых других птиц, позы, принимаемые павлином и индюком в присутствии самки, драчливость) характеризуют проявление самцового инстинкта, тогда как у самок большинства 'птиц можно отметить инстинкт насиживания и ухода за вылупившимися птенцами. Вторичные половые признаки млекопитающих частично были упомянуты выше. У некоторых видов можно отметить отчетливые различия в окраске обоих П. У антилопы нильгау (Portax p ictus) самец окрашен в голубовато-серый цвет с черной кистью на шее, а самка— в рыжий с белой полосой на груди; серый украинский скот характеризуется более темной •окраской быка; то же самое наблюдается у малайского рогатого скота (Bos banteng) (рис.5).— Отчетливый половой диморфизм имеется у человека. Мужчина отличается от женщины в среднем более высоким ростом, более толстыми костями с более интенсивно развитыми буграми в местах прикрепления мышц. (О других различиях в скелете мужчины и женщины—см. Идентификация в судебной медицине.) Подкожная жировая клетчатка у женщин развита лучше, причем отложения жира располагаются около грудных желез, на mons veneris, на бедрах и ягодицах. У готтентотских женщин стеа-топигия, по словам Дарвина, может достигать необычайной степени, затрудняющей движение.—Половой диморфизм проявляется и в устройстве гортани, которая у мужчин помимо большего объема и длины голосовых связок характеризуется более острым углом, под которым сходятся пластинки щитовидного хряща, образуя т.н. адамово яблоко. В связи с анат. особенностями гортани стоит более низкий голос мужчины по сравнению с женщиной. Тип волосяного покрова также относится к вторичным половым признакам у человека. У многих рас мужчина характеризуется наличием бороды, усов и более сильным развитием волосяного покрова на всем теле; волосы располагаются преимущественно на груди, пояснице, предплечьях и голенях. Волосяной покров на лобке и около наружных половых органов у мужчин имеет форму ромба, у женщин форму треугольника.-—П. накладывает известный отпечаток и на поведение человека, хотя констатируемые буржуазными учеными различия в психике в основном берут свое начало от уродующих воспитательных воздействий, связанных с бесправным положением женщины в капиталистических странах. Развитие признаков пола. Исследование связи вторичных половых признаков с гуморальными влияниями, распространяющимися по кровяному руслу, началось с опытов Бертоль-да (Berthold,' 1849), который удалил у петуха его семенники и пересадил их в другое место тела. Петух сохранил все характерные мужские признаки. Ряд последующих работ привел к установлению связи развития признаков П. у позвоночных с инкреторной деятельностью половых желез. Закономерность этой связи повидимому не распространяется на насекомых, т. к. опыты кастрации гусениц различных бабочек (непарного шелкопряда, капустницы) не приводили к изменениям в признаках П. Даже одновременное с кастрацией удаление за- чатков крыльев не мешало развитию как у самцов, так и у самок их типичной расцветки.— Удаление половых желез у позвоночных сказывается на их облике, причем самцы и самки разных классов реагируют на кастрацию различно. Наиболее полные данные о зависимости развития признаков пола от внутрисекреторной деятельности половых желез получены на курах (см. Кастрация). Кастрированная курица до неотличимости похожа на кастрированного петуха. На основании этих данных М. Завадовский построил свою классификацию вторичных половых признаков (см. Вторичные половые признаки). Зависимость развития некоторых половых признаков от полового гормона дала возможность «превращения пола», т. е. сообщения кастрированному самцу зависимых признаков самки и кастрированной самке зависимых признаков самца путем имплян-тации половых желез противоположного пола. Впервые эксперимент этот был , произведен Штейнахом, к-рому принадлежат термины маскулинизация и феминизация. Опыты М. Зава-довского, поставленные на курах, привели к весьма демонстративным результатам. При пересадке семенников кастрированной курице уже через 10 дней можно наблюдать усиленный рост гребня, бородок и серег и появление яркой окраски головного убора (см. отд. табл., рис. 5). Курица приобретает петуший голос и мужской половой инстинкт, короче говоря, те мужские признаки, к-рые в результате кастрации утрачиваются петухом («зависимые» или эвсексу-альные мужские признаки, по терминологии М. Завадовского) (см. отд. табл., рис. 6). Пересадка яичника кастрированному петуху (см. отд. табл., рис. 2) приводит к росту куриного оперения и к образованию гребня, по форме и окраске напоминающего куриный (см. отдельную таблицу, рис. У). Яичник, находящийся в брюшной полости, образует в большом количестве желтки, растягивающие брюшную стенку и придающие птице облик несущейся курицы. Голос такого феминизированного петуха близок к голосу курицы. Этот опыт также позволяет сделать вывод о том, что в присутствии яичника у кастрированного самца развиваются «зависимые» признаки. Общий вывод из результатов кастрации и пересадки половых желез от самца к самке и обратно сводится к тому, что ткани самца и самки в равной степени способны давать развитие как мужских, так и женских признаков. Это обстоятельство М. Завадовский обозначает как положение эк-випотенциальности соматических тканей самца и самки. Изучение влияния половых гормонов на развитие признаков П. показало своеобразное взаимодействие между мужским и женским гормоном у кур. Пересадка семенника нормальной или неполнокастрированной курице не вызывала у нее образования мужских признаков, тогда как пересадка яичника нормальному петуху, наоборот, приводила к образованию са-мочьих признаков. М. Завадовский сделал отсюда вывод о доминировании женского полового гормона над мужским. Этот вывод согласуется с результатами длительных наблюдений над кастрированными курами. Как было сказано выше, удаление яичника у курицы ведет к образованию особи с признаками, характерными для кастрата. Через несколько месяцев, без какого бы то ни было дополнительного вмешательства, у кастрированной курицы на- ч.инает отрастать петуший греоень, проявляется мужской половой инстинкт и петуший голос, происходит как бы самопроизвольное превращение в самца. Предшествующие опыты заставляют подозревать здесь наличие семенниковой ткани. Вскрытие таких кур устанавливает в брюшной полости, в области передней части почки, справа от vena cava post., округлое тело, иногда имеющее проток, который впадает в мочеточник (рис. 6). Гист. исследование этого органа указывает на то, что это мужская железа, могущая достигать полного развития, вплоть до образования сперматозоидов (рисунок 7). На основании этих опытов За-вадовский приходит к утверждению о бисексуальной природе курицы. По его данным, не только правая половая железа курицы, развивающаяся после кастрации в семенник,обладает бисексуальной потенцией, но и левая. У последней эта потенция выявляется после неполной кастрации, когда сильно поврежденная железа начинает регенерировать. В этом случае яичниковая ткань нередко частично замещается семенниковой. Курица с таким регенерирующим левым яичником нередко приобретает самцовые признаки. Петух повидимому не обладает бисексуальной потенцией, тем более, что мужской гормон,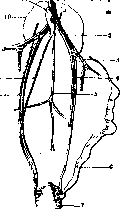
Рис.
вой 6. Положение пра-половой железы у курицы: 1—v. cava post.; ~'—v. Шаса; 3—v. magna; 4 и 5—артерии; б—редуцированный яйцевод; 7— прямая кишка; 8 — мочеточник; 9—половой проток; 20—верхняя доля почки; 11— правая половая железа.(Из Завадовского.) ,y.4«'JJ б Рисунок 7. Гистологическое строение" правой половой железы курицы. как показано выше, подавляется женским, и при наличии у петуха яичниковой ткани он неизбежно должен был бы нести признаки самки. Некоторым нарушением этой схемы являются куры т. н. куроперой расы, напр. сибрайт-бен-тамы (Sebright-Bantam), у к-рой петух не имеет ни серповидных перьев хвоста ни ланцетовидных перьев на шее и пояснице. Особенности этой породы объясняются однако не продукцией женского полового гормона мужской железой, как думал ранее Морган, а наследственными свойствами данной расы. Это выяснено перекрестными пересадками гонад от куропе-рой расы к нормальной и наоборот. Кастрированный петух сибрайт с пересаженным семенником петуха «петухоперой» расы сохраняет. куроперость, а кастрат обычной расы с пересаженным семенником сибрайта сохраняет свой тип оперения (Danforth). Результаты кастрации и пересадки половых желез у млекопитающих несколько иные. В противоположность птицам облик кастрата больше напоминает самку. Кастрированный самец антилопы нильгау приобретает рыжую самочыо окраску, самец оленя после кастрации лишен рогов; то же имеет место у баранов тех пород, где самец рогат, а самка безрога. Кастрированный мужчина лишен бороды и усов, имеет высокий голос, горизонтальный обрез волос на лобке и нередко обладает отложениями жира в тех местах, которые характерны для женщины (грудь, бедра, ягодицы) (см. Кастрация). При пересадке половых желез от одного пола к другому у морских свинок Штейнаху, Липшюцу
Рисунок 8. Сообщающиеся кровеносные сосуды хориона зародышей-близнецов коровы. (Из Завадовского.)
и др. удалось получить «превращение пола» в такой же мере, как при аналогичных опытах с курами. Есть основание предполагать, что у млекопитающих бисексуальным является самец, а самка—моносексуальна. Положение это требует однако более серьезных доказательств. Косвенным доводом в его пользу является существование т. н. «free-martin». Этим термином называется уродство развития у телят, сводящееся к тому, что наружные половые органы напоминают женские, а внутренние—ближе к мужским. Тандлер и Келлер (Tandler, Keller; 1911, 1916) и особенно Лилли (Lillie, 1917), использовавший колоссальный материал чикагских боен, показали, что «free-martin» образуется в случае разнополых двоен с общим хорионом и сообщающимися кровеносными сосудами (рис. 8) при условии, если второй партнер двойни является самцом. «Free-martin» трактуется т. о. как самка, подвергающаяся трансформации в самца под влиянием гуморальной среды своего нормального мужского близнеца. Аналогичная трансформация самки в самца была получена Бёрнсом и Вичи при сращивании попарно эмбрионов амфибий. У бесхвостых амфибий явление превращения П. наблюдается и в природных условиях, в связи с чем нередки случаи обнаружения гермафродит-ных особей (см. сводку Crew). Кастрация самца приводит к утрате ряда вторичных половых признаков и приближает облик кастрата к самке. По аналогии с млекопитающими здесь мож- но предположительно говорить о бисексуальности самца. Особенно убедительным доводом в пользу этого положения служит существование у самцов жаб т. н. Биддерова органа (см. Биддера орган), к-рый после удаления семенников превращается в нормально функционирующий яичник. Исследования Пфлюгера, Гет-вига и особенно Вичи (Pfluger, Hertwig", Witsch) показали наличие у лягушек двух рас, т. н. диференцированной и недиференцированной. У первой разделение на самцов и самок происходит еще во время метаморфоза, почему они всегда дают нормальное отношение П. У второй первоначально почти все лягушата имеют вид самок, хотя по существу являются гермафродитами, и затем около половины их б. или м. постепенно превращается в самцов. Собранный в природе материал этой расы в разное время года дает различное численное отношение полов с большим перевесом самок у молодых особей, еще не претерпевших трансформации. Наличие гормональных воздействий на развитие признаков П. у позвоночных позволило М. Завадовскому построить теорию формообразования. Эта теория сводится к представлению о процессе развития как- о реакции развивающихся тканей и органов на внешнее по отношению к ним воздействие. Образование «зависимого» полового признака протекает, по Завадовскому, по формуле х+у->А, где х—ткани, дающие признак, у—раздражитель, вызывающий его развитие, в данном случае гормон поло-бой железы, и А—готовый признак (напр. гре-вень петуха). Половой гормон, принимающий участие в формировании признаков П., произвольно выделяется из явления и как внешнее противопоставляется «реагирующим тканям». В этом сказывается формально-логическое, механистическое понимание причинности, лежащее в основе применяемого Завадовским каузально-аналитического метода. Это механистическое понимание причинности отчетливо сформулировано Миллем в основном принципе индуктивной логики—-«принципе единственного различия». Сущность этого принципа сводится к следующему. Если после введения какого-нибудь фактора появляется (или после его удаления при прочих равных условиях исчезает) известное явление [напр. исчезновение или появление признаков П. после удаления или пересадки половых желез (Бляхер)], то в таком случае этот вводимый или удаляемый нами фактор и есть причина явления. Невозможность с помощью этого принципа проникнуть в сущность явления зависит во-первых от того, что ни в одном случае нельзя быть уверенным, что вскрыто то «единственное» различие, которое определяет характер процесса, и во-вторых от того, что за решением вопроса о причине данного явления встает вопрос о причине этой причины и т. д. При этом в конце-концов приходят или к некоей первопричине, т. е. к богу, или прекращают индуктивный анализ на каком-нибудь произвольном этапе. После решения вопроса о половом гормоне как причине развития признаков пола Эванс (Evans); Завадовский и др. вынуждены поставить вопрос о причине возникновения в организме полового гормона. Соответствующая причина также отыскивается во-вне и в качестве ее признается витамин Е (витамин X), или «витамин размножения», выделенный из пророст-• ков пшеницы, листьев латука и т. д. Необходимость витамина Е для нормальной деятельности половой железы повидимому несомненна, но факт этого влияния не приближает нас к пониманию развития признаков П. Вместо проникновения вглубь явления этот метод исследования все дальше и дальше уводит нас от организма, от его специфических особенностей, изучение к-рых только и может ответить нам на вопросы о сущности явлений развития поло- вых признаков. Ведь продолжая дальше анализ, мы должны выяснить причину возникновения витамина Е в растениях, и несомненно сможем найти ее вне растения, скажем в почве. Важность и практическая ценность фактов зависимости признаков П. от эндокринных воздействий не подлежит сомнению. Необходимо только иметь в виду, что такого рода исследования не в состоянии преникнуть в сущность явления, они не вскрывают его движущих сил. его мотива, его самодвижения. Работы в этом Направлении—дело будущего. Л. Бляхер. Стимуляция полового созревания. Половая зрелость у всех животных не совпадает по времени с рождением или вылупливанием его из яйца. Она наступает как правило значительно позже. Период инфантильного—неполовозрелого—состояния у различных животных различен, но для каждого вида строго определен.В норме интервал колебания сроков полового созревания ничтожен, однако патология знает случаи сдвигов в сторону значительного сокращения. В литературе есть указания на случаи наступления половой зрелости у мальчиков и девочек в возрасте 5—6 лет (см. Pubertas ртае-еох). Во всех подобных случаях преждевременная половая зрелость стояла в связи с ранней инкреторной деятельностью половых желез. Экспериментальные исследования последних лет на разнообразных животных показали, что преждевременная половая зрелость может быть вызвана путем введения в организм инфантильного животного гипофизарного гормона. Работы Смита, Цондека, Ашгейма (Smith, Zondek, Aschheim) и др. показали, что имплянтация даже очень незначительного кусочка ткани передней доли гипофиза (до 1/1000 г весом) может вызвать преждевременное созревание половой железы. Гипофиз, взятый от человека или любого из млекопитающих животных мужского и женского П., в одинаковой степени может вызвать уже через' 100 часов после имплянтации реакцию преждевременного полового созревания у мышей. В то же время гипофиз голубей в опытах Смита давал отрицательный результат. Активной является гипофизарная железа не только от половозрелых животных, но в равной степени она активна и у инфантильных животных и даже у плода в последние месяцы беременности. До последнего времени гипофизарная железа известна как единственный раздражитель созревания половой железы, и никакая другая ткань или железа с внутренней секрецией, как показал Цондек, не может дать аналогичной реакции на половое созревание. Под влиянием гипофизарного гормона реакция полового созревания протекает бурно. В половом аппарате цнфантильного животного происходят резкие изменения как макро, так и микроскопич. порядка. Реакция полового созревания прежде всего проявляется у самок в росте и созревании фоликулов яичника, в созревании яйцеклеток, в преждевременном появлении овуляции и в преждевременном образовании желтых тел. Однако в опытах овуляция не всегда имеет место вследствие преждевременного перерождения фоликулярных клеток в лютеиновые. В этих случаях образуются т. н. атретические желтые тела, заключающие внутри себя яйцеклетку. Вследствие роста фоликулов и образования большого числа желтых тел яичник у подопытных мышей по объему и весу в 10—15 раз превосходит вес яичника от нормальных одного с ним возраста животных. Наряду с изменениями в яичнике возникают изменения также и в матке и влагалище. Матка и яйцеводы увеличиваются в объеме, из блед-ножелтых превращаются в лилово-красные вследствие сильного расширения кровеносных сосудов; в слизистой оболочке матки и влагалища происходят все характерные для течки изменения. Эпителий слизистой оболочки матки и влагалища из одно-, двуслойного превращается в многослойный, верхний слой его дегенерирует, уплощается, лишается ядер, слу-щивается и попадает во влагалищную слизь в виде безъядерных ороговелых чешуек или глы-бок. Течка, появляющаяся в период полового созревания и повторяющаяся всякий раз в период овуляции, является характерным признаком половой зрелости у всех млекопитающих животных. У человека же аналогичное явление—менструация. Течка в процессе полового созревания есть явление вторичного порядка; ее появление обусловливается не ги-пофизарным гормоном, как показал Цондек, а гормоном, продуцируемым зрелым фоликулом яичника. Трансплянтация кусочков гипофиза инфантильным или половозрелым кастратам в двойной и даже в учетверенной дозе не вызывает никаких изменений в генитальном аппарате, тогда как под влиянием фоликулярного гормона даже у кастратов появляются все характерные для течки явления. Эти опыты показали, что гипофизарный гормон оказывает влияние на половую систему только через половую железу, а через последнюю и на остальные органы генеративной системы. Созревание половой железы стоит в связи с гормоном передней доли гипофиза. В опытах вызывания полового созревания метод трансплянтации не является единственным. Большое применение имеет метод инъекции вытяжек из гипофизарной железы. Передняя доля гипофиза, какпоказал Эванс, продуцирует 2 различно действующих вещества. Одно из них стимулирует рост, а другое возбуждает половое созревание (см. Гипофиз— химия гипофиза). В силу их различной физ.-хим. природы они легко отделимы и могут быть получены из передней доли гипофиза каждое в отдельности. По своим морфогенетическим и физиол. свойствам эти два гормона являются антагонистами, что сказывается при совместном их действии. При одновременном введении в организм инфантильного животного этих гормонов преждевременное половое созревание не наступает. Следовательно гормон роста тормозит половое созревание, т. к. в отсутствие последнего во всех случаях наступает половое созревание. Выделение двух различных гормонов передней долей гипофиза Эванс пытается связать с наличием в гипофизе 2 видов железистых клеток. Базофильные клетки, располагающиеся в центральной части гипофиза, продуцируют по мнению Эванса гормон, стимулирующий половое созревание, а периферия, богатая эозинофильными клетками, выделяет гормон роста. Помимо гипофизарной железы гипофизарный гормон может быть получен также из пляценты и мочи беременных женщин (см. Про.шн).—Результаты опытов в области преждевременного полового созревания были проверены многими исследователями на различных объектах. С этой целью были испробованы амфибии, рептилии, рыбы, птицы и многие представители млекопитающих, вплоть до человекообразных обезьян. Во всех случаях удавалось стимулировать половое созреванке у самок и самцов с помощью гормона передней доли гипофиза. Гипофизарный гормон оказывает стимулирующее влияние не только на неполовозрелую -половую железу, но и на половозрелую железу, находящуюся в состоянии покоя, что наблюдается например в период спячки. В литературе имеются указания на случаи нарушения полового ритма с помощью инъекций гипофизарного гормона. В опытах с лягушками, рыбами, змеями удавалось вызвать в зимние месяца у самок яйцекладку, а у самцов половое влечение и вспышку сперматогенеза. Опыты с удалением гипофиза у хвостатых амфибий показали также, что развитие и функция половых желез стоит в связи с гипофизарной железой. В случае гипофизектомии (см. также Гипофиз, нормальная и пат. физиология) у неполовозрелых животных развитие половых желез у самцов тг самок приостанавливалось, вследствие чего не развивались и остальные органы генеративного аппарата. Яйцеводы оставались втрое уже и короче нормальных, у самцов совершенно не развивалась клоачная припухлость и затормаживалось производство зрелых половых продуктов. Все перечисленные факты указывают на то, что процесс преждевременного созревания в условиях эксперимента может быть вызван действием гормона гипофиза. Отсюда однако нельзя делать вывод, что в организме имеет место именно такая связь между деятельностью гипофиза и половым созреванием. Цондек, считающий гипофиз «мотором половой железы», основывает свое суждение на каузально-аналитическом эксперименте, с помощью которого не могут быть вскрыты связи, характеризующие организм как целое. Установление при помощи аналитического метода той или иной «причины» упрощает истинную картину взаимодействия, произвольно расчленяя организм на внешние друг другу компоненты (СМ. выше). Н. Линтварева. Определение пола. Раньше считали, что определение П. по отношению к моменту оплодотворения происходит в разное время у разных организмов, почему и различали прогамное, сингамное и метагамное определение П. Прогамное определение П. происходит до оплодотворения, сингамное—во время оплодотворения и метагамное—после оплодотворения. Однако Приходится признать, что у большинства организмов определение П, происходит в момент слияния яйца и спермия, т. е. сингамно. В наст. время можно считать доказанным, что П. каждой особи определяется известными генами, в зависимости от распределения т. н. половых хромосом. (Подробное описание аппарата половых хромосом и основанную на этом теорию определения пола Мак Кленга и Вильсона— см.Наследстжниостъ.) Теория Вильсона, согласно к-ройП. определяется половыми хромосомами, поддерживается подавляющим большинством биологов. Выдвигаемая в противовес теории Вильсона нек-рыми авторами (Hacker и др.)т. н. индекс-гипотеза, согласно к-рой гетерохромосо-мы суть не определители П., а наоборот, проявление, индекс сексуального характера особи, должна быть признана неосновательной. Защитники ее ссылаются обычно на нек-рые специальные случаи определения П., когда еще до редукции строение гамет предопределяет распределение хромосом и тем самым определение П. Эта аргументация основывается на метафизическом понимании определяющего фактора как «абсолютной первопричины», непонимании того, что 17& он также в свою очередь подвергается посторонним воздействиям как со стороны окружающих клеток организма, так и со стороны внешней среды. В вопросе о сущности генетического действия половых хромосом также имеются некоторые разногласия. Отдельные авторы (Win-ge) полагают, что ж-хромосома несет рецессивный ген женского П. (для случая мужской гетерогаметности), а у-—доминантный ген мужского П.; отсюда—хх—$, ху—&. Однако это противоречит как существованию типа хо, так и фактам, когда мужские индивидуумы вследствие уродства или экспериментального воздействия развивают известные женские признаки и обратно, из чего необходимо заключить, что и самцы и самки несут как мужские, так и женские половые гены. Поэтому Коренс (Correns) предположил, что как мужские, так и женские гены локализованы в аутосомах, в гетерохро-мосомах же находятся «реализаторы»—в ж-хромосоме ген, тормозящий проявление мужского П. или обеспечивающий возможность реализации женского, в ^-хромосоме—«тормоз» женского или «реализатор» мужского. Т. о. обходится затруднение с наличием у самцов женских генов и обратно; но существование типа хо противоречит и этой теории. Кроме того является совершенно излишним, а в методологическом отношении метафизическим усложнением, объяснять невозможность одновременного полного проявления мужских и женских генов у раздельнополых организмов существованием особых генов-реализаторов, а не характером взаимодействия между самими этими генами. Наиболее приемлемую теорию выдвинул Гольдшмидт (Goldschmidt). По его взглядам каждый гаплоидный набор аутосом заключает (для случая мужской гетерогаметности) мужской ген (или генокомплекс) М, каждая ж-хромосома—женский ген F, к-рый сильнее гена М. и каждая ^/-хромосома—качественно такой же. но более слабый, чем М, женский ген f, к-рый может и совсем отсутствовать. Соотношение силы этих генов таково, что 2F> >2M>F + f. Поэтому у особей, имеющих две ж-хромосомы, проявляется женский П., а у имеющих ху — мужской (2пхх== MMFF=$; 2пхц= MMFf=dl n—гаплоидный набор аутосом). В случае нарушения правильного соотношения между М и F (при скрещивании различных рас или вследствие присутствия лишних половых хромосом или лишних наборов аутосом) перевес женских факторов над мужскими или обратно может становиться очень малым или совсем исчезать; в этих случаях развиваются интерсексы. Интерсексы первого рода (нарушение баланса половых генов путем скрещивания рас, имеющих М и F различной силы) исследованы Гольдшмидтом на непарном шелкопряде, второго рода (Зпхх)—Бриджсом и Доброжанским на дрозофиле.В обоих случаях исследователи пришли к сходным в основном представлениям; о взглядах этих авторов дает представление таблица, взятая из статьи Бриджса (ст. 176). Считая, что каждый наследственный фактор или группа факторов обладает определенной валентностью, Бридже принимает, что «женская» валентность ж-хромосомы равна 100, а «мужская» валентность гаплоидного набора аутосом равна 80. Тогда нормальная самка, а также три- и тетраплоидные самки дадут индекс (отношение валентностей) =1,25; самец даст индекс 0,63, интерсексы—0,82. Теоретически мы- Количество дххро-мосом Колич. гаплоидных наборов аутосом Отношение валентностей Половой тип получаемой особи 3 4 3 2 1 2 1 1 2 4 3 2 1 3 2 3 1,87 1,25 1,25 1,25 l,t5 0,82 0,63 0,42 Сверхсамка ^ i тетраплоидная к j триплоидная g ' нормальная (ди-о I плоидная) ! 1 гаплоидная \ Интерсекс ! Самец Сверхсамец ! слимы, по Бриджсу, особи, у к-рых количество ж-хромосом было бы относительно слишком велико или слишком мало. Первых, имеющих индекс 1,87, Бридже называет «сверхсамками» (super-females), а вторых с индексом 0,43— «сверхсамцами» (supermales). Сверхсамки и сверхсамцы внешне мало отличаются от нормальных самок и самцов, характеризуясь пониженной жизнеспособностью и полным бесплодием. Взгляды Бриджса покоятся на цитологических наблюдениях. Оказалось далее, что интерсексуальный организм развивается до определенного момента как самец, а после—как* самка или наоборот. При этом перевес генов определенного П., недостаточно сильный, чтобы привести к образованию нормального самца или самки, влияет на время развития половых признаков: чем значительнее перевес генов данного П., тем раньше начинают развиваться присущие этому полу признаки. Общий характер этого последнего правила получил подтверждение на растительных объектах в опытах Вет-штейна (Fr. v. Wettstein) над раздельнополыми мхами, где экспериментально полученные особи FM и FFM были гермафродитны, но первые развивали антеридии в большем количестве и. раньше, чем архегонии, а вторые—наоборот. Кроме раздельнополых организмов существуют, как известно, различные формы нормального гермафродитизма (в отличие от гермафродитизма человека и раздельнополых животных, к-рый является аномалией, чаще всего интерсексуальностью). Крайняя форма нормального гермафродитизма-—т. н. пространственная м о н о й к и я, где мужская и женская половые системы функционируют одновременно у одного и того же индивидуума (кольчатые и плоские черви, многие моллюски и др.). Гольдшмидт и Вичи объясняют этот случай полным равновесием мужских и женских генов (F=M). Однако такое подведение под формулу равновесия в этом случае едва ли правильно, тем более, что и у пространственных монойки-стов мужские и женские гонады развиваются в онтогенезе очень часто неодинаково рано (см. выше о значении этого момента). Вероятнее, что здесь просто иное взаимодействие между половыми генами, при к-ром между F и М нет антагонизма, характеризующего раздельнополые организмы; т. о. F не препятствует проявлению М и обратно, а потому сравнительная сила этих генов не играет роли. Другая форма нормального гермафродитизма—к о н с е к у -т и в н а я (последовательная) м о н о й к и я, где половые системы функционируют у одного и того же индивидуума, но не одновременно, а последовательно (многие моллюски, низшие ракообразные и др.). Очевидно здесь имеется антагонистическое взаимодействие F и М, но перевес половых генов одного П. недостаточно велик, чтобы полностью исключить действие противоположного; более сильная пара генов вероятно обычно определяет предшествующую, а более слабая—последующую сексуальную фазу. Гольддшидт трактует консекутивных моной-кистов по аналогии с интерсексами как формы, у к-рых продолжительность жизни достаточно велика, чтобы охватить и мужскую и женскую фазы. У раздельнополых организмов, лишенных внутренней секреции половых желез, по его мнению естественная смерть наступает раньше «переломного момента» от одной половой фазы к другой.—Третья фаза, наиболее близкая к раздельнополости,—р удиментар-ный гермафродитизм, когда функционирует лишь одна половая система, но наряду с ней развивается временно или на всю жизнь зачаточная система органов второго П. Так, у самцов жаб имеется рудиментарный яичник (Биддеров орган), к-рый при удалении семенников может начать функционировать; у многих рас лягушек гонады самцов первоначально развиваются как яичники и лишь затем превращаются в семенники. По Вичи, у рудиментарных гермафродитов имеется относительно сильный женский фактор ^/-хромосомы—/, благодаря чему (MM—Ff) оказывается слишком малой величиной, чтобы полностью исключить развитие женских признаков; причем, понятно, чем больше f, тем значительнее рудименты гермафродитизма. У рассматриваемого типа, в отличие от двух предыдущих, имеются уже ге-терохромосомы, хотя морфологически у очень мало или совсем не отличается от ж (в исследованном случае—у бесхвостых амфибий). Вичи полагает также, что перевес мужских генов над женскими у самцов всегда должен быть по величине равен обратному перевесу у самок (FF- MM = MM- Ff). Из этого он выводит, что «сила, диференцирующая самца», спермиев, несущих ^/-хромосому, должна быть втрое больше, чем «сила, диференцирующая самок», у яиц и спермиев с ж-хромосомой, т. к. при оплодотворении яйца (с ж-хромосомой) Х-спермием баланс половых генов будет 2 (F — М), а при оплодотворении Г-спермием (M — f) — (F— M); если (M-f)-(F-M)=2 (F-M), то M-f=3(F-M). Однако вся эта вторая часть теории Вичи является произвольной попыткой подогнать все случаи определения П. под формулы равновесия. Правильной следует признать первую часть—объяснение рудиментарного гермафродитизма относительно большой величиной f. Особое место занимают два типа определения П.: однополая монойкия и прогамное определение П. Однополая монойкия (в одних случаях функциональная, в других— рудиментарная) заключается в том, что особи, генетически принадлежащие к определенному П., развиваются тем не менее как гермафродиты. Так, у круглого червя Angiostomum nigro-venosum имеется чередование поколений: раздельнополое поколение состоит из самок с 12 хромосомами(2пхх) и самцов—с 11 (2пху); спермин самцов без ж-хромосомы нежизнеспособны, и все потомство получается жж; тем не менее эти особи, генетически тождественные с самками, развиваются как гермафродиты. В сперматогенезе их одна ж-хромосома элиминируется, но спермин гермафродитов без ж-хромосомы жизнеспособны, а потому половина их потом-*ства получает 12, половина—11 хромосом; возобновляется раздельнополая генерация. Рудиментарная однополая монойкия наблюдается напр. у веснянки (Perla marginata), у к-рой самцы имеют морфологически хорошо развитый, но неспособный к функционированию яичник. Отличие этого последнего случая от рудиментарного гермафродитизма амфибий состоит в его бесспорно вторичном характере. Происхождение однополых монойкистов от раздельнополых форм не подлежит сомнению как вследствие полной раздельнополости почти всех остальных представителей тех классов, к к-рьш они принадлежат, так и вследствие наличия у них совершенно отчетливо развитого гетеро-хромосомного аппарата (у Angiostomum и у Per 1а—хо). Как упомянуто выше, в нек-рых случаях (у нек-рых червей и насекомых) распределение половых хромосом предопределяется строением половых !клеток еще до редукции. Так, у фи-локсеры существуют две категории партено-генетических самок: из яиц одной категории выходят только самцы, из другой—только самки; у червя Dinophilus apatris неоплод отворенные яйца бывают двух резко различных сортов: крупные и мелкие, причем из первых выходят только самки, из вторых—только самцы; у цек-рых тлей, принадлежащих к типу ХО, перед редукционным делением весь хондриом (см. Митохондрии) скопляется у одного полюса, и ж-хромосома всегда отходит именно к этому полюсу. Это—случаи т. н. прогамного определения П. Во всех этих случаях хотя распределение хромосом и подвергается воздействию посторонних факторов, но определение П. в точности следует этому распределению. Т. о. прогамное определение П. не опровергает, а подтверждает хромосомную теорию определения П. Что же касается возможности воздействия внешних факторов на хромосомный аппарат, то такая возможность a priori существует и во всех остальных случаях сингамного определения П.; на распределение половых хромосом и тем самым на П. потомства удается воздействовать и экспериментально (t° и т. д.; важнейшая работа в этом направлении принадлежит Seiler'y). Препятствием для хромосомной теории это может казаться лишь при метафизическом непонимании роли определяющего фактора и проблемы взаимодействия. Характер воздействия половых генов на морфогенез половых признаков не может еще считаться вполне выясненным. По Гольдшмидту,. все вообще гены суть проферменты, под влиянием к-рых в развивающемся организме образуются гормоны, к-рые уже непосредственно определяют формирование отдельных признаков организма. Однако это пока только гипотеза. Установлен твердо лишь различный характер действия половых факторов у позвоночных и беспозвоночных. У этих последних развитие вторичных половых признаков совершенно независимо от половых желез. Филогенетическая эволюция П. и определяющего его механизма очевидно шла от гермафродитизма к раздельнополости. Особенно ясно это видно у растений, где раздельнополы в обычном смысле только нек-рые представители высших цветковых (раздельнополость нек-рых мхов имеет совершенно особый характер и она также представляет собой вторичное явление). Но и у животных гермафродитно подавляющее большинство низших, а раздельнополы почти все высшие формы. Наиболее примитивной является очевидно пространственная монойкия, при которой, по нашему представлению, имеются две пары генов (или генокомплексов): FFMM, причем одна пара не препятствует проявлению другой, а потому сравнительная сила их безразлична. При возникновении антагонизма между F и М мужская и женская.половые системы уже. не могут развиваться одновременно, и пространственная монойкия превращается в консекутивную. F и М вообще говоря не равны; если в одном из генов более сильной пары (скажем F) произойдет мутация (трансгена-ция), существенно ослабляющая его, то FFMM превратится в FfMM. Т. о. наряду с особями, гомозиготными по обоим половым генам (самками), окажутся особи, гетерозиготные по женскому фактору (самцы). Хромосома, несущая f, имеющаяся только у одного П. в одном экземпляре, есть ^-хромосома, т. о. мы получаем законченный механизм определения П. у раздельнополых организмов. Пока f сравнительно велико, несущие его организмы являются рудиментарными гермафродитами; при дальнейшем его уменьшении (или исчезновении) они становятся вполне раздельнополыми.-—Целый ряд факторов заставляет принять, что ^-хромосома и ген f в ходе эволюции деградируют; сюда относятся установленные факты отсутствия генов в ^/-хромосоме, так что наличие нескольких у у одной особи не оказывало никакого влияния на ее фенотип (в том числе и на половые признаки—следовательно здесь / = о); почти всегда меньшая величина у по сравнению с х, наконец—существование типа хо. Тот факт, что у ряда высокоорганизованных животных, в частности у человека, ?/-хромосома имеется налицо, в то время как ее нет у многих насекомых и червей, не противоречит общему деградирующему ходу ее эволюции, т.к. раздельнополость несомненно независимо возникала многократно в различных пунктах эволюционного процесса, и деградация ^/-хромосомы конечно шла неодинаково быстро в различных ветвях филогенетического дерева. Изложенная гипотеза в нек-рых существенных пунктах отличается от взглядов Вичи, также признающего примитивность пространственных монойкистов, но полагающего, что существование у них мужской и женской системы объясняется равновесием половых генов (F = M). Но т. к. у раздельнополых организмов F yMy>f {при женской гетерогаметности), то Вичи вынужден принять, что F и / изменяются всегда одновременно и в противоположных направлениях, причем f всегда уменьшается на величину втрое большую, чем увеличение F [т. к. по Вичи (M-f)=3(F- M)—см. выше]. Такая натянутая конструкция необходимо вытекает из подведения под схему равновесия в проблеме пространственной монойкии и в вопросе о сравнительной величине генного баланса у самцов и у самок (FF— ММ и ММ - Ff). Вместе с тем Вичи не учитывает возможности различного взаимодействия между F и М у раздельнополых и гермафродитных организмов (проблема антагонизма). На эту возможность Гольдшмидт • указал еще в 1920 г., но не сделал отсюда неизбежного вывода, что у пространственных монойкистов нет необходимости принимать равновесие F и М. Стремление к универсализации схемы равновесия у наиболее крупных исследователей проблемы определения П. является ■естественным следствием отсутствия у них сознательно применяемого диалектического метода. Половой д и морф из м—см. Диморфизм. В. Вепдровский. Лит.: Гольдшмидт Р., Механизм и физиология определения пола, М.—П., 1923; Завадовский М., Пол и развитие его признаков, М., 1922; о н ж е, Пол животных и его превращение, М.—П., 1923; он hi е, Динамика развития организма, М., 1931; Морган Т., Теория гена, Ленинград, 1927; Синнот Э. иДенн Л., Курс генетики, М.—Л.. 1931;Филипченко Ю., Общий курс генетики, Москва—Ленинград, 1928; о н ж е, Экспериментальная зоология, Ленинград—Москва, 1932; Hartmann M., Verteilung, Bestimmung und Yererbung des Geschlechts bei Protisten u. Thallophy-ten, В., 1929; L i"p s с h ii t z J., Die Pubertatsdriisen und ihre Wirkungen, Bern, 1919; Meisenheimer J., Geschlecht und Geschlechter 1m Tierreich, B. I, Jena, 1921; W i t s с h i E., Bestimmung und Vererbung des Geschlechts bei Tieren, В., 1929. Смотрите также:
Смотрите также:
- ПОЛЕ ЗРЕНИЯ представляет собой пространство, все точки которого одновременно видны при неподвижном взгляде. При монокулярной фиксации изображение фиксируемого объекта получается в области желтого пятна, изображения же предметов, расположенных на нек-ром расстоянии от ...
- ПОЛЕВАЯ САНИТАРИЯ, прикладная отрасль военной гигиены, охватывающая собой систему профилактических мероприятий по улучшению и оздоровлению условий пребывания войск в полевой обстановке в целях охраны здоровья бойца и повышения его боеспособности. П. ...
- ПОЛЕВАЯ. ХИРУРГИЯ, учение о мед. помощи раненым. Термин «полевая хирургия» введен. Пироговым. Предметом П. х. является изучение действия на человеческий организм механических средств вооруженной борьбы и организация хир. помощи пострадавшим ...
- ПОЛЗАНИЕ, вид физич. упражнений, передвижение по земле с помощью нижних и верхних конечностей. Значение П. для целей физ. развития — ограниченное, оно используется лишь в нек-рых системах гимнастики в качестве ...
- ПОЛИАДЕНИТ (polyadenitis), множественное воспаление лимф, желез. Наблюдается при острых и хрон. инфекционных заболеваниях как регионарно, напр. в области шеи (корь, скарлатина, дифтерия), средостения (пневмония), так и повсеместно (сифилитический, туб. П.). П. ...