ПЕРЕКРЕСТ ХРОМОСОМ
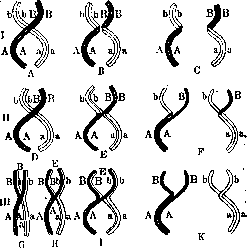
Рисунок 1. Схема хиазматипии: I—схема одновременного обмена обеих нитей каждой из сложных хромосом («перекрест четырех нитей»); II—схема обмена одной из нитей каждой из сложных хромосом («перекрест двух нитей»); III—интерпретация наблюдаемых цитологических картин при отрицании гипотезы хиазматипии.
мыми генами не абсолютно, то они могут попасть в известном проценте в разные гаметы; это происходит благодаря обмену между гомологичными хромосомами, происходящему до редукционного деления. Этот обмен и называется перекрестом хромосом, или крос-синговером-(crossing-over). Изучение процентов обмена между многими генами одной и той же хромосомы обнаружило следующую закономерность: «если а, о и с обозначают три гена и если отношение сцепления а с & и Ъ с с известно, то отношение а к с является функцией суммы или разности ab и be». Так (у дрозофилы), гены желтого цвета тела и белых глаз, находясь совместно в хромосоме, попадают в разные гаметы в 1,2%, гены же белых глаз и вильчатых жилок—в 3,5%. Если же исследовать процент перекреста между геном желтого цвета тела и геном вильчатых жилок, то получается 4,7%, т. е. ровно сумма полученных ранее значений. Стертевант-(Sturtevant) (1913) и отчасти Мел л ер (Ми Пег) установили, что эта закономерность объясняется линейным расположением генов в хромосоме и что процент перекреста есть функция расстояния. В виду этого возможно построить карту хромосомы, т. е. выразить линейно относительные расстояния генов. Процент перекреста между двумя генами может иметь любое значение от 0% до 50%, т. к. при 50% наблюдается уже независимое распределение генов. Между тем суммирование процентов перекреста всех линейно расположенных в хромосомах генов дает обычно величины большие 50 и даже 100. Это объясняется явлением т. н. двойного перекреста: если мы наблюдаем обмен группы генов- b- , то в случае одновременного перекреста между А и Ъ и Ъ и С получатся гаметы такого типа, как будто между in Сне происходило никакого обмена: АЬСиаВс (рис. 2). Поэтому если гены А я С настолько удалены друг от друга, что между ними могут происходить двойные перекресты, то процент попадания А и С в разные гаметы не будет равен сумме процентов А+В+С, а меньше последней на величину процента двойных перекрестов. Двойные перекресты могут проис-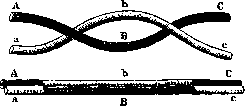
Рисунок 2. Схема двойного перекреста хромосом.
анверсии части хромосомы на 180°. Все эти факты говорят за то, что процентом перекреста как функциям расстояния нельзя придавать абсолютного значения. Действительно, Меллер и Пейнтер (1928) при цитологическом исследовании случаев выпадения (deletion) большей части хромосомы обнаружили, что хромосома, генетически равная приблизительно лишь Via части целой, цитологически оказалась равной около 1/4 неповрежденной хромосомы. Наконец последние данные Доб-жанского (Th. Dobzhansky, 1929, 1930) также показали, что, если в отношении расстояния между генами генетические данные не совсем совпадают с цитологическими, то линейная последовательность генов оказалась блестяще подтвержденной (рис. 3). Цитологические процессы, лежащие в основе П. х., в настоящее время следует считать невыясненными. Гипотеза хиазмапатии Янсенса, поддержанная Морганом, по к-рой на одном из стадий перед редукционным делением хромосомы попарно перекрещиваются и обвиваются друг вокруг друга, разрываются в местах перекрестов, а затем разорванные участки соединяются, обменявшись друг с другом,—признается большинством цитологов недоказанной (рис. 1). Зейлер (Seiler, 1922) выдвинул другую теорию П. х., по к-рой хромосомы на ранних стадиях состоят из отдельных свободных участ-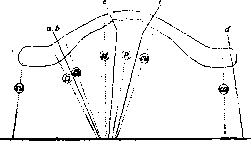
Рисунок 3. Сравнение генетического (нижняя горизонтальная линия) и цитологического (вверху) плана расположения генов в третьей хромосоме Drosophila melanogaster на основании изучения экспериментальных отрывов частей хромосомы под влиянием Х-лучей. Пунктиром указаны сравнительные места расположения генов в обоих планах; а, Ь, с, d и е—места экспериментальных разрывов хромосомы,- ru—rougaoid; D—Dichaete; th—thread; si—scarlet; p—peach; cu—curled; ca—claret.
ков, свободно комбинирующихся, а затем ассоциирующихся в целые хромосомы. Но невыясненность цитологических основ П. х. никоим образом не колеблет современных генетических представлений в области хромосомной теории наследственности. Лит.: Морган Т., Структурные основы наследственности, гл. IV—XII, М.—П., 1924; Bridges С. a. Morgan Т., The third-chromosome group of mutant characters of Drosophila melanogaster, p. 9—30, Washington, 1923 (методика изучения перекреста хромосом); М u 11 е г Н., The mechanism of crossing-over, Am. natur., v. L, 1916,- Morgan Т., В r i d g e s C. a. Sturter v h ant A., Genetics of Drosophila, chapt. X, S.-Gravenage, 1925; Seiler J., Die Crossing-over Studien der Schule Morgan, Naturwissenschaften, 1924, д» 34 А. Гайсинович. Смотрите также:
Смотрите также:
- ПЕРЕКРЕСТНАЯ ИММУНИЗАЦИЯ, способ иммунизации, имеющий целью выяснить иммунобиологические взаимоотношения двух или более микробов. Если перекрестная иммунизация производится микробами А и Б, то одна группа животных иммунизируется культурой микроба А, а другая ...
- ПЕРЕЛИВАНИЕ КРОВИ, см. Трансфузия крови.
- ПЕРЕЛОМЫ, всякое полное нарушение целости твердого предмета (Wegner), в данном случае кости. П., являясь результатом наиболее тяжелых травм, составляют одну из самых серьезных глав травматологии. По статистике Брунса (London Hospital ...
- ПЕРЕМЕЖАЮЩИЙСЯ, интермиттирующий (от лат. intermittere—оставлять промежуток). П. характер б-ни может зависеть от разных причин: 1) от цикла развития паразита и от хода иммунизаторных реакций (ремиссии неновые приступы при малярии—febris intermittens ...
- ПЕРЕНОСЧИКИ, живые посредники в деле распространения инфекций и инвазий. Распространение возбудителей инвазий и инфекций в большом количестве случаев происходит при прямом или косвенном посредничестве животных. Последние получают возбудителя из места ...