ОБЛИТЕРАЦИЯ
Рисунок 4. Влияние на рост крыс прибавления недостающих аминокислот.
аминокислот делает их полноценными (рисунок 4). В. Лавров. Пуриновый обмен (физиология)— см. Мочевая кислота, Нуклеопротеиды, Пургтовые основания. Патология белкового обмена. Кюличест-венныенаруш-ения белкового обмена бывают при голодании полном и" частичном и при кормлении неполноценными'белками. Повышение белкового обмена (т. н. токсический распад белка) наблюдается при нек-рых отравлениях (пантопоном, окисью углерода, сулемой, щавелевой к-той), при злокачественных опухолях, при некоторых формах б-ни Базедова, при диабете, особенно экспериментальном, при острых инфекционных заболеваниях, при лихорадочных процессах. Обратное явление'—усиленное отложение белков в организме (Eiweissmast)— положительный баланс азота—легче всего достигается после периода голодания, при хрон. недостаточном питании и особенно легко в период выздоровления после истощающих болезней. Задержка N в организме имеет место даже при сравнительно небольших количествах вводимого белка. При усиленной мышечной работе происходит отложение белка вследствие развития рабочей гипертрофии мышц, сопровождающейся часто значительной задержкой азота в организме. Вопрос о возможности достигнуть отложения белков в нормальном состоянии у взрослого организма подвергался большим дискуссиям. Решение этого вопроса имеет большое практическое значение особенно в животноводстве. Кельнером (Kellner) было доказано, что при рациональном кормлении у коров удается достигнуть значительного отложения белков в организме. У человека также-можно получить отложение белковых веществ, учитываемое по положительному азотистому балансу. Эта задержка N в организме достигается как усиленным введением белковых веществ, так и увеличением (кало-рически) количества безазотистых пищевых продуктов, предохраняющих часть белков от энергетического расходования. Задержка азота в организме не может с достоверностью рассматриваться только как процесс отложения белков; тем не менее большие количества задержанного N (Grafe) не могут зависеть лишь от азотистых шлаков, но являются также в результате накопления белковых веществ, хотя образование их и не сопровождается обычной задержкой воды в организме. В виду несоответствия в количестве задержанного N и воды большинство авторов считает, что этот образованный белок не тождествен с белком протоплазмы, но частью циркулирует в соках организма (Voit) частью откладывается в печени (Rub-ner, Noorden) или в клетках др. органов (Lutje, Berg). Чтобы проверить, является ли отложившийся белок биологически таким же, как и белок протоплазмы, исследовался основной обмен, который однако при задержке N не дал повышения. Это наблюдение говорит против образования истинного белка, являющегося составной частью протоплазмы, как это бывает в случаях отложения белка при рабочей гипертрофии и после истощения. Представляет ли такое отложение белков какое-либо преимущество для организма, трудно сказать. Некоторые авторы считают отложение белков полезным бла-« годаря тому, что эти белки вследствие сво-* ей лябильности предохраняют белки протоплазмы от распада и кроме того в случае необходимости могут превращаться в истин-s ные белки. Качественные изменения белкового обмена. Кроме физиол. бе:;т ков органов и жидкостей при болезненных процессах находят особые белки, вероятна происходящие путем перестраивания тканевого белка, а именно: амилоид (см. Амилог идное перерождение) и белок Бене-Джонса (см. Бене-Джонса белковое тело). Бене-Джон-совский белок появляется в моче обычно при множественных миеломах, но не во всех случаях этого заболевания. Иногда в ткани миеломы выпадают кристаллы Бене-Джон* совского белка (Вульф). Бенс-Джонсовский белок был кроме того обнаруживаем в моче при остеомаляции, саркоматозе, гипернефроме, лейкемии. Бенс-Джонсовский белок б. ч. выделяется одновременно с сывороточным белком; больная почка легче пропускает Бенс-Джонсовский белок, чем здоровая. Место перегруппировки белка неизвестно; по-*. видимому клетки костного мозга влияют на это перестраивание белковой молекулы. Готшальк (Gottschalk) проводит аналогию между Бенс-Джонсовской альбуминурией и диабетом; в обоих случаях существует нарушение межуточного обмена с образованием вещества.частично усваиваемого клетками организма. Другие авторы проводят параллель между образованием пигментными опухолями меланогена и меланина и образованием Бенс-Джонсовского тела в миеломах. При впрыскивании Бенс-Джонсовского белка развивается поражение почек (Krauss). Под влиянием парентерального введения Бенс-Джонсовского белка развиваются процессы иммунизации. Изменения в ассимиляции аминокислот являются важными нарушениями белкового обмена. При заболеваниях печени наблюдаются гипераминацидурия (при острой атрофии печени кристаллы лейцина и тирозина) наряду с гиперацидемией. Повышенное выделение аминокислот с мочей бывает при многих инфекционных б-нях, диабете, лейкемии. Существуют также заболевания, характеризующиеся недостаточн. межуточным усвоением одной или нескольких аминокислот.— Цистинурия или цистиновый диатез (Wollaston) характеризуется повышенным выделением с мочой i-цистина в растворенном состоянии или в виде осадка; иногда цистин дает в пузыре или почках камни. Сущность цистинурии состоит в том, что организм не в состоянии расщепить молекулу цистина. Повидимому цистинурия является нарушением межуточного обмена, обусловленным общим недостаточным дезаминиро-ванием в виду того, что обычно наряду с ци-стином наблюдаются аномалии распада других аминокислот (лейцина, тирозина, аспара-гина) и аминов (кадаверина, путресцина). Появление при цистинурии диаминов представляет конституциональную аномалию и отличается по своему значению от диамину-рии, вызываемой путем действия бактерий в кишечнике на лизин и аргинин, дающие при распаде те же амины.—Алкаптонурия (см.) как б-нь не имеет большого практич. значения, но в теоретическом отношении сыграла большую роль при изучении распада арома-тич. веществ в человеческом организме. Нуклеиновый обмен. Физиол.-хим. исследования обмена нуклеопротеидов в организме и пат. нарушения этого обмена имеют большое значение для выяснения патогенеза подагры (см.). Для подагры характерно отложение моноурата натрия в хрящах, сухожилиях (tophi); в крови содержание мочевой к-ты б. ч. повышено, в то же время выделение эндогенной мочевой к-ты мочой понижено; экзогенная мочевая к-та выделяется также недостаточно и замедленно. А. Б. Геррод, Тангаузер и Лихтвиц(А. В. Garrod, Thannhauser, Lichtwitz) считают, что явления подагры зависят от первичной недостаточности почек в отношении выделения мочевой кислоты. Бругш и Шиттен-гельм (Brugsch, Schittenhelm) объясняют подагру повышенным содержанием в организме человека мочевой к-ты, вызванным недостаточными ферментативными процессами, преимущественно понижением активности или отсутствием уриколитического фермента, разрушающего в норме мочевую к-ту, к-рая образуется в процессе межуточного обмена (см. Мочевая кислота, Подагра). Креа-тинурия—см. Креатин. Вопрос об образовании и нахождении а м-м и а к а в организме за последние годы подвергся коренному переем отру. Нашем и Бенедиктом найдено, что главным местом образования аммиака являются почки. Количество аммиака, выделяемого с мочой, во много раз превышает количество аммиака, циркулирующего в крови. То же самое подтверждено Амбаром и Шмидом (Ambard, Schmid). В крови аммиак находится в* двух формах (Parnas): во-первых в очень незначительном количестве (0,015 мг % N) в виде нона аммония, во-вторых в виде аденин-ну- клеотида (2 мг % N)—«основное аммиачное вещество» Парнаса (Ammoniakmuttersub-stanz—AMS), к-рое легко отщепляет аммиак при стоянии сыворотки. В почках этот ну-клеотид расщепляется при кислой реакции, образуя аммиак. Образование аммиака тесно связано с нейтрализацией к-т, если только они в организме не сгорают до С02. Из соотношения между выделением аммиака и наличностью кислот выясняется влияние питания на выделение ЫЩ. При мясной пище выделяется много аммиака потому, что из белков образуется много неорганических кислот (серная к-та, фосфорная к-та); при смешанной пище аммиака меньше, при растительной пище совсем мало. При оральном или парентеральном введении к-т количество аммиака в моче увеличивается. Одновременно с этим в сыворотке количество ионизированного аммония не изменяется, но ясно уменьшается содержание неионизированной части (AMS). То же самое наблюдается и при эндогенном ацидозе. Наибольшие повышения в выделении аммиака наблюдаются при диабете: количество аммиака доходит до 12 з в сутки вместо нормальных количеств 0,5—1 г. При введении щелочей не наблюдается изменений в обеих аммиачных фракциях крови, но резко понижается количество аммиака, выводимого^ мочой. При поражении почек, особенно при нефрите, выделение аммиака понижено. При уремии, несмотря на состояние ацидоза, при сильном понижении резервной щелочности количество аммиака в моче резко понижено; в крови в то же время содержание аммиака не повышается. Гассельбах (Hasselbalch), указавший на значение NH* как регулятора нейтрализации, отметил, что при нормальных условиях у одного и того же лица существует определенное соотношение между аммиачным числом (отношение азота аммиака к общему количеству азота) и концентрацией ионов водорода; у почечных б-ных этот коеф. меньше, т. к. больная почка или совершенно не в состоянии образовать аммиак или не может образовать его в достаточном количестве. При введении к-ты нефритику аммиак в моче совсем не повышается или повышается лишь на несколько сантиграммов в противоположность здоровому, у которого содержание аммиака повышается на 1—1 У2 г В СУТКИ. Ю. Гефтор. Пат. анатомию нарушений белкового обмена—см. Амилоидное перерождение, Белковое перерождение, Гиалиновое переро-оюдение, Кахексия, Маразм, Мутное набухание, Подагра, Уремия. V. Жировой обмен. Жировой обмен в широком смысле слова— совокупность явлений, представляющих собой судьбу «жира» в организме. Сюда относятся следующие главнейшие моменты: проникновение пищевого жира из кишечника в кровь, транспорт жира в ткани, его потребление и откладывание в тканях, синтез жира. В понятие «жир» может быть включено и понятие о жироподобных телах, или т. н. липоидах; т. о. в жировом обмене рассматривается: I. Собственно жировой обмен; II. Липоидный обмен. 70в I. Собственно жир ов ой о бмен. Из кишечного канала после всасывания (см. Пищеварение) жиры проникают в организм двумя путями: 1) через лимфатич. капиляры ворсинки, образуя в них род тонкой эмульсии, отчего лимфа в более крупных лимф. сосудах брыжейки просвечивает в периоды всасывания как белая масса; обогащенная .жировыми капельками лимфа (chylus) через ductus thoracicus вливается в венозн. кровь; по нек-рым наблюдениям (например Munk, Friedenthai) над девушкой с фистулой duc-tus thoracici главная масса жира переходит в организм именно этим путем. 2) Через капиляры ворсинки и далее через воротную вену в кровеносную систему печени и т. д. По наблюдениям Эррико (Errico) кровь, взятая одновременно из воротной и шейной вен животных, находящихся на высоте пищеварительного процесса (жировая пища), имела содержание жира в воротной вене ■большее, чем в ше7!ной, чего не могло бы быть, если бы жир переходил в венозную систему исключительно через ductus thoracicus. Отсюда следует, что жир переходит в кровь обоими путями. Появление жира пищи в лимфе начинается уже в первый час пищеварения, и спустя два часа содержание жира в хилусе ductus thorac. доходит до 3—14% (смотря по содержанию жира в пище и скорости всасывания) против нескольких pro mille в лимфе натощак. Содержание жира в крови в часы всасывания пищи также 'постепенно возрастает (алиментарная гипер-. липемия). Так наз. «нормальная» липемия, т. е. содержание жира натощак (после предварительного «среднего» смешанного режима питания или после свободного от жира первого завтрака у человека), у разных животных и даже у разных индивидуумов одного и того же вида дает очень значительные колебания—бт 0,2% до 0,8% (у нек-рых авторов показаны значительно более низкие числа). В качестве иллюстрации распределения жира (жирных к-т) в крови приводится таблица Гориуши (Horiuchi). Табл. 13. С р а в н и т е л ь н о е распределение жирных к-т в крови. Человек и животные л и и й ° ё 5*5 Мужчина............0,36 Женщина............ 0,38 Собака..............j 0,52 Кролик (без жировой диеты) . \ 0,28 Кролик (жировая диета) . . . I 0,34 0,38 0,40 0,22 0,30 0,36 0,29 0,36 0,42 Величина алиментарной липемии подвержена многим, частью случайным влияниям (скорость всасывания и количество пищи, количество в ней жира, его усвояемость, активность ферментов, состояние эндокринной системы и т. д.). Иногда плазма крови на высоте пищеварения делается непрозрачной вследствие накопления в ней яшровых шариков. В качестве примера для липемии приводятся наблюдения Блура (Bloor) на собаке. Через 24 часа после еды жир крови равнялся 0,6%, через Зх/4 часа после дачи еды 100 ем3 оливкового масла—0,73%, через 6V4 час. после дачи 100 см* оливкового масла— 1,20% и через 8 час. после дачи 100 см* оливкового масла—0,87%. У человека алиментарная липемия возникает еще легче, чем у собаки, между тем как у кролика она обнаруживается с трудом. У гусей при откорме (при наличии жира в пище) алиментарная липемия достигает 3% в крови и до 6% в сыворотке. При обильном углеводном кормлении, когда углеводы превращаются в жир, липемии не наблюдается, конечно при отсутствии в пище жира (Bleibtreu, Rosenf eld). Понижение содержания жиров в крови до нормы является следствием того, что жировые частицы «улавливаются» во-первых жировой тканью, где жир скопляется в клетках как запасный материал; такими тканями («жировыми депо») являются подкожная клетчатка, межмышечная соединительная ткань, сальник и др.; во-вторых жировые частицы могут перейти в другие ткани, где жир потребляется в энергетических или структурных целях.—При целом ряде пат. состояний (см. ниже патологию), при голодании, при экспериментальном панкреатическом или флоридзиновом диабете наступает также липемия, причем жир в этом случае поступает в кровь из жировых депо организма; такая липемия совпадает с потерей запасов гликогена в организме, и мобилизация жира здесь может быть рассматриваема как приспособление организма к новым условиям сильно пониженного углеводного обмена. Пример такой липемии приведен в работе Мишера (Miescher) над голодающим лососем (биол. голодание—см. Голодание). Из всего известного нам о липемии т. о. явствует, что в организме могут возникать такие условия, при к-рых капилярн. система становится проницаемой для жира то в ту то в другую сторону. Механизм такой установки организма совершенно не известен. Нек-рые авторы пытались рассматривать исчезновение жира из крови после гиперлипемии как следствие его расщепления липазой крови. Однако, если такой липолиз и имеет место, все же повидимому он не является безусловно необходимым для перехода жира из крови, т. к. интравенозно введенный окрашенный жир переносит краску в те ткани (печень, костный мозг), куда он обычно транспортируется (Biondi, Neumann). Исчезновение жира из крови нек-рые исследователи пытаются объяснить исходя из того предположения, что жирные к-тьт и жир дают соединения с белками плазмы, к-рые могут выходить . за ппеделы кровеносной системы. В пользу возможности такого соединения жира с белком говорят следующие факты: 1) трудность извлечения (количественного) всего жира крови путем экстракции жировыми растворителями без предварительного применения пепсин-соляной к-ты для растворения белков; 2) белковый раствор в виде напр. трансуцатной (асцитной) жидкости после смешивания его с определеннымколичеством масла не целиком отдает последнее экстрагирующему жир растворителю: в белковом растворе остается еще некоторое количество масла, переходящее снова в растворитель только после действия пепсин-соляной к-ты. К такому же «маскированию* жира ведет и действие на него желчных кислот в крови, в результате к-рого также получаются растворимые в плазме соединения. На «маски рова-ние» жира указывает также и работа Бион-ди-Неймана (Biondi, Neumann), которые изучали ультрамикроскопическую картину крови человека; кровь, взятая во время всасывания жира, обнаруживала в темном поле многочисленные блестящие капельки (гемо-конии), причем количество таких частиц по мере всасывания жира постепенно увеличивалось, достигало некоторого максимума (через 6 часов после приема пищи), сноЕа падало и часов через 12 доходило до нуля. В пробах крови, взятых у человека натощак, отсутствовали гемоконии. Можно думать, что мелкие капельки жира могут проходить через стенку капиляров. Следующие цифры (Mockel) иллюстрируют в процентах концентрацию жира в тканях жирного животного (собака): кожа—38%, подкожн. слой—87%, мышцы—19,6%, печень—13,4%, мозг—-12,7%, кости—12,0%. В периоды обеднения печени гликогеном жир обильно переходит в печень из жировых депо, поднимая в ней содержание жира до 30—40%.—Введение в организм жира (так же как и введение углеводов и белков) вызывает на некоторое время повышение обмена (специфическое динамическое действие) (см. белковый обмен). Однако такое повышение в сравнении с действием углеводов и белков незначительно, что объясняется вероятно легким переходом жира в жировые депо. В опыте Магнуса-Леви (Magnus-Levy) при даче человеку 210 г сливочного масла увеличение обмена против основного составляло 9—14% в течение седьмого часа и 6—8%— на восьмом часу. Внешняя температура оказывает очень существенное влияние на проявление специфического динамического действия жиров. У Рубнера имеется следующий пример: собака получала 171,3 кал. жира на 1 кг веса. Табл. 14; Внешняя te Теплопродукция на 1 кг веса (в калориях)
Здоровый человек (68 кг) содержит 12,36 кг жира (приблизительно 18% своего веса). У откормленных животных жир доходит до 50% их живою веса. "Что жиры пищи при значительном их потреблении транспортируются в жировые депо, видно из целого ряда опытов. Так, при кормлении собак и мышей окрашенным свиным салом краска переходит во все жировые депо, остаются неокрашенными только печень и надпочечники; краска переходит также и в молочный жир (Jacobsthal); очевидно, что откладывание
жира в печень является вторичным процессом—туда жир транспортируется из других тканей; впрочем при очень обильном введении жира окрашивается и печоноиный жир. Кроме того установлено, что в жировых депо может откладываться «чужеродный» жир, если он вводится с пищей длительное время и в большом количестве; так, собаки после продолжительного кормления бараньим салом накопляют его в своей подкожной клетчатке. То же самое отмечено при кормлении бараньим салом золотых рыбок и карпов. Травоядные, потребляющие зеленый корм, имеют жир более твердый, чем при зерновом корме (по Rosenfeld'y жир зеленых кормов—твердый, жир злаков—жидкий). Испытаны были на животных различные жиры—репное масло, льняное, се-самовое, коровье, кокосовое и бараний жир—и все они могли быть обнаружены не только в жировых депо подопытных животных, но и в молоке, в куриных яйцах и в копчиковой железе птиц. С другой стороны, каждый вид животных обладает своим собственным, характерным для него жиром. Это нисколько не противоречит вышесказанному, поскольку каждый вид имеет и характерную для него комбинацию пищевых средств. Наконец большие количества жира возникают не из жиров, а из углеводов пищи или из белков. Промежуточный обмен ж и р о в. Жирные к-ты при своем горении в организме претерпевают постепенное «ступенчатое» разрушение, для которого является характерг ным окисление /З-углеродного атома, причем в этом месте происходит разрыв цепи и образование карбонильной группы. СНа — СН2 — СНа — СНа — СН2 — СООН сна — сн2 — сн2 — со -/- сн2 — соон СН2 — СН2 — СНа — СООН СН2 —СО Ч- СИ, — СООН сн2 — соон. Очевидно, что при каждой фазе окисления отщепляются два атома С (в виде уксусной к-ты или ее дериватов); очевидно также, что к-ты с четным числом С-атомов должны перейти через стадий четырехуглеродной цепи—масляной к-ты; к-ты с нечетным числом С-атомов масляной к-ты не образуют; наоборот, одним из последних промежуточных продуктов у них является пропионовая к-та. Опыты на животных подтверждают и уточняют эту схему разрушения жирных к-т в организме. Так, Кнооп (Кпоор), воспользовавшись наблюдениями Ненцкого, Саль-ковского и др., показавшими трудность сгорания бензольных ядер в организме, вводил в организм фенилдериваты, имеющие различные жирные боковые цепи, к-рые не успевали сгорать в теле до конца, останавливаясь на нек-рых промежуточных продуктах, переходящих в мочу. Таким способом было выяснено, что жирные к-ты действительно подчиняются общему закону очередт ного окисления Д-углеродов и последовательного отщепления от цепи двууглеродных отрезков. Так, Д-фенилпропионовая кислота 7 09 СН2—СН2СООН переходила в бензой- ную к-ту 1 ICOOH, между тем как фенил- уксусная I \ СН2СООН и миндальная к-та СН(ОН)—СООН выделялись из организ- ма без изменения (у них не было в боковой цепи /?-атома для окисления); фенилмасля- ная к-та СНЙ—СН2—СНЙ—СООН и ее /3-й у-дериваты давали фенилуксусную к-ту. Отклонения от этого закона имеются, но они не противоречат ему и объясняются специальными хим. условиями; так напр. жирные цепи с аминовой группой в а-положении перед окислением дез'аминируются, и дальнейшее окисление, согласно общему правилу, идет в этом пункте цепи (т. е. в а-С). Различное отношение организма к к-там с четным и нечетным числом' С-атомов было показано Эмбденом на переживающей печени, а именно, жирные к-ты, будучи прибавлены к жидкости, проводимой через печень, дают ацетон, причем ацетонообразова-тельные свойства у кислот с четным числом углеродных атомов значительно выше, чем у к-т с нечетным числом; кроме того обнаружено, что с удлинением цепи к-ты уменьшаются ацетонообразовательные свойства. Табл. 15. гво вше-тона л Вещества, прибавленные к пропускае- с "5 щ мой через чпечень крови gSri«H Кол] обра гося в мг кров Норм, маслян. к-та СН3(СН2)2С00Н » валериан. » СН3(СН2)3С00Н » капронов. » сн3(сн2)4соон юо » гептилов. » СН3(СН2)5С00Н i 12 » октиловая » СН3(СН2)6С00Н » нониловая » СН3(СН2)7СООН » декановая » СН3(СН2)8С00Н Интересно здесь же отметить, что к-ты с нечетным числом С-атомов с прямой цепью при прохождении через переживающую печень не только являются слабыми ацетоно-образователями, но и угнетают ацетонообразовательные свойства к-т с четным числом С, если вместе с ними прибавлены к жидкости, пропускаемой через печень (антикето-ногенное действие). Опыты школы Эмбдена показали, что жирные к-ты с четным числом С-атомов при прохождении через переживающую печень служат материалом для образования ' ацетоуксусной к-ты. Весь этот ряд фактов позволяет сделать вывод, что жирные к-ты с четным числом углеродных атомов проходят при своем разрушении через стадий масляной к-ты, к-ты с нечетным числом—через стадий пропионовой к-ты. Оба эти промежуточных продукта подчиняются общему закону окисления у Д-углеродов. Так, масляная к-та должна превращаться в /?-оксимасляную к-ту или ацетоуксусную к-ту, пропионовая (аналогично фенилпро-пионовой к-те)—в /5-океипропионовую или /?-альдегидопропионовую к-ту и далее СН8—СН2—СНз-СОО Н -> СН3СН2СООН СНз—СО.СЩ—СООН И а ,СН3—СНОН.СН2—СООН г*8 СН20Н-0Н2—СООН СН—СНаСООН в ацетальдегид (дальнейшая судьба этого соединения см. углеводный и белковый обмен). Что касается вообще самого механизма окисления /S-углерода, то согласно теории Виланда (Wieland) появлению кетокис-лоты предшествует образование ненасыщенного соединения с одной двойной связью (дегидрир ование) н н R—С—С—СООН ! ! К, — С = С — СООН. Н Н дегидрирование Ф) (а) Доказательством наличности этого процесса служат опыты Лизиса и Гартли (Leathes, Hartley), которые показали, что при введении в организм животных фенилпропионовой к-ты L J—СН2—СН2—СООН в моче можно получить фенил-коричную кислоту II СН — -СН.СООН, а Декин кроме того при таких же условиях нашел /8-фенил-/3-оксипропис~ новую к-ту и /?-фенил-/?-кетопропионовую СНОН—СНЙ—СООН и о- со— к-ту —СН2—СООН и соответственно ацетофенон и бензойную к-ту. Подводя итог этим и др. исследованиям, Декин считает, что предельные к-ты через стадий непредельной к-ты переходят далее в окси- и кетокислоты, разрываясь затем на месте кетонной группы. Общая схема окисления представляется следующим образом: н н н н -Н20 | | +Н20 I | -НаО К—С— С—СООН-э-R—С=С—СООН-»К—С—С—СООН-* • I I • II Н Н ОН Н (8 а 4-Н20 ->R—Ct^CH2-COOH->R—COOH+CHs—СООН. !i о Слабым местом схемы по Декину является превращение оксикислоты в кетокислоту. Океикислоты в организме вообще используются с нек-рым трудом, и правильнее представить себе, что оксикислота появляется как побочная реакция восстановления кетокислоты; действительно при скармливании бензоилуксусной к-ты (0-фенил-кетопропи-оновая кислота) С6Н5.СО.СН2.СООН была получена соответствующая спиртокислота Z-C6H5CHOH.CH2COOH. Кроме того инъекция голодающему животному под кожу /J-оксимасляной к-ты не увеличивает выде- •23 ления ацетоуксусной кис лотыи ацетона.но такое увеличение имеет место после инъекции масляной к-ты. С другой стороны, известно, что/?-оксимасляная кислота в изолированной печени легко окисляется в ацетоуксусную кислоту. Наиболее верным поэтому может считаться такой взгляд (Dakin), что все окси-, кето- и ненасыщенные кислоты находятся между собой в состоянии подвижного равновесия и могут при этом переходить одна в другую. При отрыве двухуглеродного отрезка у /^-углеродного атома у последнего возникает карбоксильная группа, отрываемый же отрезок представляет собой вероятно уксусную к-ту, к-рая может общим порядком или окисляться до С02 и Н20 или перейти через янтарную (получаемую в результате синтеза 2молекул уксусной к-ты), фумаровую, яблочную, щавелево-уксусную и пировиноградную к-ты в молочную кислоту и далее участвовать в образовании углеводов. В пользу превращения жиров в углеводы в организме говорят недавние работы Шатерникова и Молчановой по изучению поглощения кислорода и выделения углекислоты жировой тканью на голодающем баране (путем сравнения газообмена до и после ампутации значительной части жирового хвоста). Выяснилось, что дыхательный коефициент (см.) у жировой ткани 0,34—0,35; теоретический коеф. Шово (Chauveau), данный им в его формуле перехода жира в углеводы: 2C57HiioOe + 6702 = 16СвН120„ + 18С02 + 1Ш20 равен Ц = 0,268. II. Липойдный обмен. Вопрос об обмене липоидов (см.) прежде всего сталкивается со сложностью самого понятия; т. к. в это понятие объединен целый ряд химически различных групп и соединений [фос-фатиды, стерины (напр. холестерин), цере-брозиды и др.], то при рассмотрении обмена приходится отдельно разбирать эти группы. Физиология очень бедна сведениями по обмену липоидов вообще; наиболее изученным телом липоидной природы является относящийся к группе фосфатидов лецитин (см.). Под влиянием ферментативной работы кишечного канала лецитиноподобные вещества могут распадаться на свои составные части: так. лецитин дает холин, фосфорную кислоту и глицерин. Все авторы отмечают быстроту всасывания лецитина; местом этого всасывания повидимому является тонкая кишка, однако до сих пор неразрешенным является вопрос о том, необходимо ли для всасывания лецитина предварительное его расщепление или нет. Неизвестны также с точностью пути продвижения лецитина и друг, за пределы кишечника; из опытов Эйхгольца (Eichholtz) видно, что после дачи больших количеств лецитина содержание его в крови поднимается на незначительную величину; можно предположить поэтому, что лецитин идет через v. portae в печень, где и задерживается; по крайней мере из нек-рых опытов видно, что кровь v. portae богаче лецитином, чем кровь периферических вен; Сальковский также указывал, что печень накопляет лецитиноподобные тела после их обильного введения в кишечник; различные органы неодинаково относятся к приему лецитинов: в то время напр. как печень накопляет их в большом количестве, другие ткани, напр. мозг, ничего подобного не обнаруживают. Из кишечника фосфатиды всасываются также и лимф. путями: Словцову удалось показать увеличение липоидного фосфора и азота в лимфе через 5 часов после дачи лецитина.—Вопрос о синтезе фосфатидов в животном организме был поставлен очень давно и разрешался по-разному; выводы из опытных данных сделать весьма трудно. Здесь можно только остановиться на исследованиях Мек Коллема и др. (McCollum), показавших, что куры на пище, практически лишенной лецитина (снятое молоко и рисовая мука), продуцировали 294,5 г желтка с 36,51 г фосфатидов, из которых 8,83 г приходилось на лецитин. Подобное же наблюдение над утками было сделано Фингерлингом(Р^ег1^), в опытах которого эти птицы синтезировали лецитин за счет неорганического фосфора пиши. Обмен липоидов группы стеринов — см. Стерины, Холестерин. Вопрос о всасывании стеринов пищи в кишечнике, при своем разрешении наталкивается на большое затруднение вследствие того, что много холестерина выводится из организма с желчью, секретами кишок, отторгнутым эпителием. Часть холестерина, попавшего в кишку из пищи и желчи,всасывается,часть разрушается бактериями, превращаясь в другие стерины (копростерины). На собаках (14—15 кг веса) с, фистулой желчного протока было установлено выделение холестерина в желчи 0,04—0,08 г, содержание же этого вещества в кале у контрольной собаки было 0,21—0,32 г pro die. Холестерин пищи всасывается в количествах до 54 % (у собак) .Прибавка жира, увеличивает всасывание холестерина, прибавка углеводов ухудшает всасывание его. При значительной прибавке холестерина к мясу заметное количество его исчезало из кишечника (почти 38%). Однако до сих пор остается неясным, можно ли исчезновение холестерина из кишечника считать только выражением его усвоения. Фюрт (О. Fiirth) обращает внимание на возможность частичного бактериального разрушения холестерина в кишечнике. Далее было замечено, что увеличение белка в пище (а не холестерина в ней) вызывает увеличение холестерина в желчи. Интересно также отметить, что распад эритроцитов, вызванный введением в организм толуилендиамина, увеличивает выделение холестерина с желчью. На стационарных желчных фистулах кроме того выяснено, что увеличение дачи холестерина (с пищей) не отражается непосредственно на содержании его в желчи, но вызывает (временно) повышение холестеринемии, причем, когда последняя достигает нек-рого б. или м. высокого уровня, начинает увеличиваться выделение холестеринажелчью. Механизм перехода холестерина через стенку кишечника неясен; в некоторых опытах было установлено увеличение холестерина в хилусе (например Munk-Rosenstein'a на девочке с хи-лусной фистулой) вместе с увеличением жира в пище. Высказывалось соображение, что холестерин «двигается по тем же путям, как и жир». Вопрос о синтезе холестерина в организме решается положительно (наблюдения над содержанием холестерина в куриных яйцах до и во время их насиживания). У младенцев при исключительно молочном' питании выделение холестерина превышает поступление его с пищей раза в 1у2—ЗУа, между тем количество холестерина при росте нарастает; это видно из следующих таблиц 16 и 17. Табл. 16. Холестерин Фактор Пища пищи кала 540 » » женск. . . 1200 » » » . . 0,050 0,072 0,107 0,070 0,125 0,114 0,212 0,177 0,125 0,424 2,3 2,9 1,7 1,8 3,4 а б л. 17. Возраст ев « А <° я И rt £ о к о. и в Возраст ев В СО Е-1 ей [-£0 О A i ar R rt КОЮ ребенка о о о о, Я» ев ребенка rt<U с? а> » о я £ о н£ ] Ufflo Новорожд. 3 050 1,507 9 м. . 6 000 7,682 Ц м. . . 3 800 4,272 18 м. . 9 000 12,140 5 м. . . 5 700 5,883 Если при росте количество холестерина абсолютно возрастает везде в тканях, как и в мозгу, то при наличии постоянного отрицательного баланса можно предполагать, что в теле где-то идет его новообразование. По отношению ко взрослым в более новых исследованиях также подчеркивается, вопреки старым данным, превышение выхода холестерина над поступлением. Поэтому и у взрослого можно предполагать новообразование холестерина в теле. Разрушение холестерина в организме мало еще выяснено; во всяком случае это тело является очень резистентным. У голодающих собак, потерявших до 2/3 своего веса и очень значительную часть своих жировых депо, общее содержание холестерина в теле оказалось очень близким к таковому контрольных животных; это показывает, что холестерин не разрушается в теле при голодании (Beumer) и отношение холестерина к высшим жирн. кислотам сильно сдвигается в сторону первого. Интересно отметить, что отношение холестерина к фосфатидам остается без изменения. Из организма холестерин удаляется не только через желчь, но частично и через кожу. В секрете сальных желез холестерин всегда присутствует и у некоторых" животных выделяется этим путем в больших количествах. Так, у барана выделяется ланолин, содержащий очень много сложного эфира высших жирных кислот и холестерина. Из других стеринов наиболее изученным в последнее время является так называемый эргостерин. (Происхождение жиров из углеводов—см. обмен углеводный; происхождение жиров из белков^—см. Белковый обмен). б. Лавров. III. Липоидно-углеводный обмен. В то время как переход углеводов организма в жиры является общепризнанным фактом (см. углеводный обмен), обратный переход жиров в углеводы еще очень многими авторами оспаривается (Тангаузер). Имеются лишь косвенные доказательства возможности перехода жиров в углеводы через стадий кетонов. Еще Шово подчеркивал роль жиров в энергетических процессах организма. Он считал, что необходимая для организма энергия доставляется непосредственным сгоранием углеводов, причем это могут быть и углеводы, произошедшие из белков и жиров. Эта предпосылка все более и более подтверждается наблюдениями над процессами обмена при разных пат. и фи-зиол. состояниях (см. жировой обмен). Во главе сторонников теории гликонеогенеза из жиров, причем не только при патологических, но и при физиол* условиях, стоит Геельмюй-ден (Geelmuyden). Изучение липемии, гли-козурии.кетонурии, величины D:N, дыхательного коефициента при различных состояниях (диабет, гликозурия беременных, кетону-рия при неукротимой рвоте новорожденных, фосфорн. и флоридзиновое отравление, апан-креатический диабет, голодание, анемия, физ. напряжение и т. д.), исследования по вопросу о судьбе целого ряда кетогенных и анти-кетогенных субстанций заставляют признать, что углеводный и жировой обмены сопряжены между собой. Промежуточным продуктом при переходе жиров в углеводы являются кетоновые тела. Магнус-Леви показал, что кетоны, образующиеся при распаде белка при coma diabeticum, ни в какой мере не покрывают количества кетонов, выводимых мочой, следовательно роль жиров в образовании кетонов не должна вызывать сомнений. Переход жиров через ацетоновые тела в углеводы при пат. процессах также почти общепризнан. Геельмюйден же считает, что кетоновые тела являются физиол. продуктом обмена и что в норме тоже происходит, правда очень незначительный, гли-конеогенез из жиров. Кетоновые тела в незначительном количестве циркулируют в крови и только в том случае, если они не успевают в организме перейти в гликоген, они накопляются в избытке и выводятся почками. Эмбден и его сотрудники доказали, что печень является единственным органом, где образуются кетоновые тела, дальнейшая же их переработка может протекать как в печени, так и в других тканях. Количество кетонов, подвергающихся полному превращению в печени, зависит от наличия в ней антикетогенных субстанций (нек-рые аминокислоты, глюкоза и гл. обр. гликоген). При недостатке антикетогенных субстанций в печени кетоновые тела переносятся кровью на периферию, где при наличии необходимых условий подвергаются окончательному метаморфозу. Переход жиров в углеводы протекает повидимому либо путем ^-окисления (Кнооп) либо по Лизису (Leathes), т. е. путем дегидрирования, а затем уже /5-окис-ления. Регуляция липоидно-углевод-ного обмена со стороны нервной системы осуществляется повидимому из дна III желудочка, гл. обр. infundibulum и tuber cinereum, верхним шейным сегментом спинного мозга и п. splanchnicus. В гормональной регуляции главная роль принадлежит инсулину и адреналину (Вертгеймер, Рааб и др.); менее ясна роль гормона средней доли гипофиза. Открытие инсулина, регулирующего углеводный обмен, способствующего переходу углеводов в жиры и так резко влияющего на липоидный и кетоновый обмены, дало новые доказательства реальности гликонеогенеза из жиров. Вертгеймер считает, что инсулин является катализатором при переходе жиров и кетонов в углеводы, причем в первую фазу его действия в углеводы переходят лишь жиры, находящиеся в печени. Под влиянием больших доз инсулина гликоген исчезает, под влиянием малых вновь образуется (влияние то на образование жиров то на образование углеводов). Под влиянием инъекций адреналина новообразование гликогена из жира ускоряется и усиливается, при этом азотистый распад не увеличивается, гликоген печени нарастает, а жир исчезает. Ряд экспериментальных работ свидетельствует также, хотя и менее убедительно, о возможности новообразования сахара из жиров под влиянием питуитрина. с. Жиелнн. Патология жирового обмена. Жиры доставляются животному организму отчасти в готовом виде, отчасти образуются в обмене из других веществ и биологически играют роль резервов, вступающих в обмен при разнообразных условиях и могущих значительно влиять на общий обмен своей высокой калорийностью. Важное значение жира видно также из того, что в здоровом состоянии всякий организм содержит довольно много жира, а при хронич. заболеваниях в большинстве случаев наблюдается похудание, зависящее гл. образ, от исчезновения жира. Только крайне редко жир практически исчезает совеем, обычно все же небольшое его количество может быть обнаружено в жировых депо. Причина исчезновения жира далеко не всегда ясна; иногда похудание может зависеть от недостатка вводимой пи-Щи (отказ от еды, невозможность глотать, непроходимость привратника и т. п.). К исчезновению уже отложенных жиров ведет также недостаточное всасывание жира пищи. Расщепление и всасывание жиров из кишечника страдают при недостатке ферментов поджелудочной железы и отсутствии желчи! В этих случаях часто наблюдаются сильные поносы, к-рые еще более нарушают питание. Это же токсическое или кахекти-ческое исчезновение жира имеет место при большинстве истощающих заболеваний. Исчезновение жира обнаруживается раньше всего путем взвешиваний больного и при клиническом осмотре. Гораздо труднее потерю жира выявить контролем обмена веществ, т. к. образование и распад жиров представляют медленно текущие процессы, требующие долгосрочных наблюдений, а образующиеся продукты обмена для учета нуждаются в применении сложной методики в неудобной для б-ного форме (газовый обмен). К этому присоединяется еще то обстоятельство, что исчезновение жира сопрово- ждается также и потерей, а отложение—задержкой воды в тканях (до 40% в жировой ткани). При недостаточности отложения жира, равно как и при избыточных жировых накоплениях, последовательность процессов образования или расщепления жиров не страдает, жировой "обмен в своих отдельных этапах от нормального не отличается, и нигде в организме не получается образования межуточных продуктов обмена, равно как не отмечается появления таковых в выдыхаемом воздухе или в моче. Точных измерений, позволяющих определенно говорить о чрезмерном или недостаточном отложении жира, не существует, и все вопросы, касающиеся необходимости откормить больного или уменьшить его вес, решаются на основании всей клинической картины. Отмечается, что есть люди, которые остаются худыми, несмотря на безусловное переедание. Неспособность пополнить жировые запасы нек-рыми авторами даже выделяется в особую болезненную форму, к-рая ставится в связь с расстройством желез внутренней секреции. Чаще однако приходится иметь дело с пациентами, склонными к'полноте. И здесь расстройства внутренней секреции имеют существенное значение, но в чем состоит механизм повышенного отложения жира, не совсем еще ясно. Промежуточные продукты жирового обмена—ацето-уксусная и /?-оксимасляная к-ты в норме исчезают почти без следа, что объясняется их окончательным окислением. В случаях пат. ацидоза значительная часть ацетоновых тел *не окисляется, а выделяется в мочу. Причина, вызывающая чрезмерно большое поступление кетонообразующих веществ в кровь, состоит в обеднении печени гликогеном, вслед за чем наступает сильнейшая мобилизация жирных к-т, к-рая и ведет к образованию оксимасляной и ацетоуксусной к-т. Т. о. развитие кетонурии и кетонемии наступает как вторичное явление после исчезновения гликогена из печоночной клетки; первоначально наступает расстройство обмена углеводов. Антагонизм между отложением в печени гликогена и наполнением печоночной клетки жиром наблюдается кроме различных гликозурий также и при всевозможных отравлениях (фосфор, мышьяк, хлороформ и др.), под влиянием бактериальных токсинов, особенно также при периодической рвоте детей.—Среди группы липоидов наиболее характерными особенностями обладает холестерин, обмен которого изучен лучше других. Болезненные явления, связанные с холестерином, можно различать как повышенное содержание коллоидального холестерина или как отложения этого вещества в нерастворенном состоянии. Вероятно существуют также изменения, касающиеся отношения холестериновых эстеров к свободному холестерину (см. Холестерин). Нарушение холестеринового обмена с накоП' лением холестерина в организме и отложе нием его в тканях ставят в связь с происхождением атеросклероза артерий и желчнокаменной б-ни. Действительно, при этих заболеваниях (а также при сахарном диабете и иногда при хрон. нефрозе) находят увеличение содержания холестерина в крови. Экспериментально удается, вводя долгое время холестерин, вызвать у некоторых животных (преимущественно у кроликов и • свинок) отложения его в стенках сосудов, очень напоминающие картину атеросклероза человека. е« Фромгольд. Пат. анатомия нарушений жирового обмена в смысле избыточного отложения нейтрального жира—см. Оысирение. Отложзние холестерина обычно в результате гиперхо-лестеринемии проявляется в артериосклерозе (см.), в картинах ксантелязм и псев-доксантом (см. Кеантома), в образовании холестериновых желчных камней. Своеобразную пат.-анат. картину дают нек-рые специальные заболевания, связанные с нарушением жирового обмена вещзств и отложением жира и липоидов в тканях (см. Гоше болезнь, Ниман-Пша болезнь). Пат. анатомия местного расстройства жирового обмена клеток—см. Жировое перерождение. VІ. Углеводный обмен. Поступающие с пищей полисахариды (крахмал, гликоген) и дисахариды подвергаются гидролитическому расщеплению под влиянием пищеварительных ферментов до стадия моносахаридов, которые, как и пред-образованные моносахариды, поступившие с пищей, всасываются главным образом в тонких кишках. В нормальных условиях дисахариды, не подвергшиеся .гидролизу, хотя они и растворимы в воде, не всасываются. Поступающая с растительной пищей клетчатка (целлюлеза) подвергается лишь частичному гидролизу под влиянием кишечных бактерий.—Судьба глюкозы после всасывания. Если проследить за содержанием глюкозы в крови воротной и печеночной вен, то мы увидим, что содержание глюкозы в крови педоночной вены во время пищеварения мало меняется; в воротной же вене содержание ее во время пищеварения резко увеличивается; на него влияет также количество углеводов в пище. Такая разница в содержании глюкозы в двух вышена-. званных венах объясняется тем, что избыток глюкозы, поступающей в воротную вену во время всасывания, задерживается в печени, отлагаясь в ее клетках в виде гликогена. Превращение глюкозы в клетках печени в гликоген есть процесс ферментативный. Гликоген, отложенный в клетках печени, является запасным углеводом; под влиянием имеющихся в печени ферментов этот гликоген может снова расщепиться на молекулы глюкозы, к-рая переходит в кровь. В печени человека может отложиться до 150 з гликогена. Такое же приблизительно количество его м. б. отложено в мышцах. Кроме того гликоген в небольших количествах может отлагаться почтив каждой клетке тела животных. Если в пище слишком много ди- и моносахаридов, то по воротной вене к печени приносится сразу очень большое количество глюкозы и часть глюкозы, не превратившись в печени в гликоген, проходит дальше в пе-чоночную вену, что вызывает повышение содержания глюкозы в сосудах большого круга кровообращения, т. е. наступает т. н. пищевая (алиментарная) гипергликемия избыток сахара из крови выделяется почками, наступает т. н. пищевая гликозурия. Пищевые гипергликемия и гликозурия прекращаются, как только уменьшается доставка простых углеводов с пищей. Различные углеводы с неодинаковой легкостью превращаются в печени в гликоген и откладываются в ней; так например ассимиляционная способность печзни для глюкозы равна 100 г, т. е. у здорового человека гликозурия наступает при введении per os сразу больше 100 з глюкозы. Ассимиляционная способность печени по отношению к галактозе равна только 40 з. При болезнях печени, особенно при заболеваниях печоночной паренхимы, ассимиляционная способность печени уменьшается. Поэтому определением ассимиляционной способности пользуются для фнкц. диагностики печени. Углеводы откладываются прозапас не только в виде гликогена, но могут также превращаться в жиры, которые откладываются в жировой ткани. Во время голодания гликоген, отложенный в печени, превращается в глюкозу. Последняя переходит из печени в кровь, а из крови поступает в различные ткани, где и сгорает. Гликоген и мышечная работа. При работе мышц содержание гликогена в них, а также в печени уменьшается. Это было впервые установлено Клод Бернаром (Claude Bernard), нашедшим, что у зимне-спящих животных гликогена много в мышцах и в печени; когда же они, проснувшись после зимней спячки, начинали двигаться, содержание гликогена и в мышцах и в печени уменьшалось. Кюльц(Кй1г) нашел,чтов теле собаки, возившей в течение десяти часов тележку с грузом, было около 1 згликогена на 1 кг веса тела, в то время как в теле других собак, не проделавших такой работы, было около 38 з гликогена на 1 кг веса. Т. о. при работе мышц тратятся углеводы; если работа короткая, то уменьшается содержание гликогена в мышцах. При более продолжительной и тяжелой работе уменьшаются запасы гликогена также и в печени. Дыхательный коеф. при интенсивной физ. работе равняется единице. Промежуточный обмен углеводов в мышцах. Превращения углеводов в мышцах, связанные с освобождением энергии, проходят через ряд промежуточных этапов, приводящих к образованию конечных продуктов окисления углеводов, угольной к-ты и воды. Прежде всего гликоген, имеющийся в мышцах, расщепляется на молекулы глюкозы. Если гликогена в мышцах нет, то глюкоза поступает в мышцы из крови. Затем глюкоза вступает в соединение с фосфорной к-той, и образуется глюкозомонофос-форная к-та (см. Лактацидоген). Этот процесс происходит под влиянием фермента фосфатазы. Лактацидоген расщепляется на фосфорную кислоту и активную (лябильную) форму глюкозы, легко подвергающуюся дальнейшим превращениям; расщепление лактацидогена происходит под влиянием фермента гексозомонофосфатазы. Следующим этапом обмена углеводов является превращение активной глюкозы в 2 частицы метил-глиоксаля, причем возможно, что в качестве промежуточного продукта образуется сперва метилглиоксальальдоль. Фермент, катализирующий этот процесс, называют глико-лазой. СН2ОН СН2 СНа СН3 ! СНОН СОН СОН или СО I -НОН I I | СНОН-------> СНОН—» СОН СОН I -НОН I СНОН СН СНа СНз I! СОН или СО СОН СОН СОН СОН глюкоза метилглио- метилглиоксаль ксапьальцоль (2 частицы) Метилглиоксаль далее под влиянием ке-тон-альдегидмутазы (или глиоксалазы) превращается в молочную кислоту, к-рая, подвергаясь дегидрированию, превращается потом в пировиноградную к-ту: СН» СН« СНя I ! -Н2 [ СО + На—* СНОН —> СО I 1| ■ I I СОН О COOH СООН молочная пировиноград-к-та ная к-та Пировиноградная к-та может образоваться прямо из метилглиоксаля в результате дегидрирования его гидрата под влиянием оксидоредуказы (дегидразы): СНз -Но | СНз СНд +Н20 сон со н с—он ^он со I соон Пир овинограднаякислота под влиянием открытого Нейбергом фермента карбоксилазы, катализирующего отщепление от пировино-градной к-ты частицы С02, превращается в уксусный альдегид.Уксусный альдегид окисляется затем в уксусную к-ту, а эта последняя окисляется в конце-концов до С02 и Н20. сн I со сн. +HOH сн сон ~ уксусный I альдегид CООН С02 3-Hj ,Н СН, с^он соон \птт УксУсная к-та ^ОН гидрат уксусного альде- Таков же ход превращения углеводов при спиртовом брожении (см.), изучение к-рого помогло во многом выяснению обмена углеводов в животном организме; только при спиртовом брожении уксусный альдегид не окисляется в уксусную к-ту, а восстанавливается в этиловый спирт. При спиртовом брожении гексоза также сперва вступает в соединение с фосфорной к-той, причем образуется гексозодифосфорная к-та. Образование некоторых из вышеуказанных промежуточных продуктов (напр. метилглиоксаля, уксусного альдегида) доказано экспериментально (Neuberg). Зимазу (см.), представляющую собой комплекс ферментов, обусловливающих процессы как спиртового брожения, так и гликолиза, можно разделить на апозимазу и козимазу. Одни этапы обмена углеводов протекают только при наличии обоих компонентов зимазы, другие же могут иметь место и при наличии одной апозимазы; так напр. превращение гекеозы в метилглиоксаль может итти в отсутствии козимазы (при наличии только-апогликолазы); дальнейшее же превращение метилглиоксаля в молочную к-ту может итти только при наличии кофермента (при наличии апоглиоксалазы и коглиоксалазы). Ней-берг удалял козимазу или ослаблял ее действие; тогда он обнаруживал накопление метилглиоксаля, т. к. в отсутствии кофермента его дальнейшее превращение не имело места* Образование уксусного альдегида Ней-берг доказал с помощью метода улавливания уксусного альдегида путем связывания его сульфитом натрия или кальция. Если; обмен углеводов протекает в присутствии сульфита, то уксусный альдегид, образуясь-из цировиноградной к-ты, вступает с ним в соединение; такой связанный уксусный альдегид не может подвергаться дальнейшим превращениям, и он накапливается в качестве конечного продукта углеводного обмена (спиртового брожения); его можно выделить и количественно определить. При брожении окисление метилглиоксаля в пировиноградную к-ту и восстановление уксусного альдегида в спирт являются двумя противоположными, между собой связанными процессами: уксусный альдегид восстанавливается при помощи того водорода, который освобождается при окислении (дегидрировании) гидрата метилглиоксаля. При брожении в присутствии сульфита восстановление уксусного альдегида невозможно, и тогда водородом, освобождающимся при дегидрировании метилглиоксаля, восстанавливается другая его частица: в результате образуется глицерин, к-рый вместе с уксусным альдегидом (связанным с сульфатом натрия) и с С02 являются конечными продуктами этой формы спиртового брожения. Уксусная к-та при своем окислении проходит повидимому через следующие этапы:. в= результате дегидрирования двух ее молекул образуется янтарная к-та, к-рая при дальнейшем дегидрировании превращается в ненасыщенную фумаровую кислоту; последняя, присоединяя воду, превращается в яблочную к-ту, к-рая путем дегидрирования превращается в щавелево-уксусную. Из щаве-лево-уксусной в результате отщепления СОа, образуется пировиноградная к-та, от к-рой отщепляется затем еще одна молекула С02, и образуется уксусный альдегид, окисляющийся затем в уксусную к-ту. Процесс идет дальше таким же порядком, так что молекулы уксусной к-ты постепенно одна за другой окисляются до С02 и воды (водород, который освобождается при дегидрировании вышеуказанных промежуточных продуктов,. окисляется кислородом до воды). Процессы углеводного обмена являются обратимыми реакциями: они могут итти в обоих направлениях—как в сторону расщепления и образования все более простых продуктов, так и в сторону синтеза глюкозы из промежуточных продуктов ее распада. Регуляция углеводного обме-н а. В углеводном обмене огромную роль играют два гормона: адреналин и инсулин. Адреналин и инсулин оказывают влияние на гликогенную функцию печени и они являются в этом отношении антагонистами::. 722: адреналин стимулирует процессы распада гликогена в печени, иначе говоря, превращение его в глюкозу; адреналин кроме того усиливает проницаемость почек для сахара. Инсулин стимулирует синтез гликогена в печени из углеводов, если они есть, или из белка или жира в случае углеводного или полного голодания; инсулин стимулирует также переход глюкозы из крови в ткани и окисление углеводов в тканях. Вследствие такой роли этих гормонов введение в организм адреналина вызывает гипергликемию и гликозурию и сопровождается уменьшением содержания лактацидогена в мышцах. Введение инсулина вызывает, наоборот, гипогликемию; при введении больших доз инсулина, вызывающих сильное обеднение тканей углеводами, могут наступить гипогли-кемические судороги. Инсулин, как это показали исследования Нейберга и Готшалка (Gottschalk), а потом Альгрена (Ahlgren), усиливает образование уксусного альдегида в тканях. По мнению некоторых авторов (Винтер, Смит) инсулин стимулирует превращение глюкозы крови в активную форму глюкозы (так называемую гамма-глюкозу). Угле|водный обмен и витамины. Расстройства углеводного обмена наблюдаются также при отсутствии витаминов в пище. Функ указал, что экспериментальный полиневрит наступает тем скорей, чем больше углеводов в пище голубей, и что во время полиневрита содержание сахара в крови изменяется. Подробно изучена связь между витаминами и углеводным обменом для экспериментальной цынги (скорбута). Установлено (А. Палладии), что при скорбуте, вызванном отсутствием в пище витамина С, кривая сахара крови испытывает определенные изменения: сперва она повышается (иногда после небольшого кратковременного понижения) и достигает определенного максимума, после к-рого начинает уменьшаться; наконец гипергликемия сменяется гипогликемией, достепенно увеличивающейся вплоть до смерти животного. Появление клин, симптомов цынги (кровоизлияние в деснах, шатание зубов, характерный запах изо рта и т. д.) совпадает с переходом гипергликемии в гипогликемию, т. е. с моментом/когда содержание сахара в крови оказывается близким к норме. Поэтому при исследовании содержания сахара в крови у людей, т. е. при первом появлении клин, симптомов цынги, находят его близким к норме. Параллельно с изменением кривой сахара крови меняется и содержание диастазы в ней. Содержание гликогена в печени по мере развития цынги постепенно уменьшается, и наконец печень оказывается лишенной гликогена. Такова уже в общем картина изменений кривой сахара в крови и при экспериментальном полиневрите (Collazo), обусловленном отсутствием в пище птиц витамина В, и при питании пищей, лишенной всех витаминов. Таким образом при отсутствии витаминов в пище животных организм утрачивает способность использовать нормальным образом углеводы. Авитаминоз в отношении расстройств углеводного обмена напоминает диабет, тем более, что введение инсулина при авитаминозе уменьшает гипергликемию, так же как уменьшает гипер гликемию при диабете. А. Палладии. Патология углеводного обмена. Расстройства углеводного обмена встречаются нередко. У постели б-ного они могут не сопровождаться никакими характерными симптомами, и для обнаружения их нужны анализы мочи, дыхательного обмена, исследование крови и т. д. Учитывая, что в нормальной моче при обычных условиях питания можно. открыть лишь следы глюкозы, которые при перекармливании сахарами возрастают до количеств, точно определяемых, можно установить предельное число граммов моно- и дисахаридов, которое не дает такого повышения нормальной гликозурии и к-рое характеризует толерантность. Отмечается повышение толерантности в случаях dystro-phia adiposo-genitalis и при микседеме, когда б-ные иногда переносили огромные количества углеводов (500 г глюкозы). Чаще можно* встретить обратное явление—пониженную толерантность, к-рая характерна для диабетического обмена. Понятие «толерантность» следует отличать от понятия «усвояемость» (ассимиляция), под к-рой разумеется количество углеводов, вступивших в обмен (разрушенных или задержанных). Если здоровому давать глюкозу в количестве, превышающем толерантность, то не все, что дано сверх толерантности, выделится с мочой, и чем больше глюкозы давать, тем больше ее будет выделяться, но тем больше будет и потребляться организмом. Нарушение толерантности и гликозурия иногда наблюдаются и не при сахарной б-ни (отравления, апоплексия, инфекции, болезни печени). Различные виды сахара вступают в обмен и разрушаются неодинаково легко. Фруктоза и галактоза напр. особенцо легко выделяются мочой при заболеваниях печени. Галактоза обнаружена также в моче грудных детей при расстройстве кишечника. Наконец описаны случаи появления в моче фруктозы и различных пентоз. Случаи чистой фруктозурии, т. е. появление в моче фруктозы без одновременной гликозурии и при условии отсутствия плодового сахара в пище,— довольно редки. Они протекают, как легкий диабет, с к-рым могут быть наследственно-связаны. Наблюдения пентозурии интересны тем, что иногда пентоза выделяется в оптически недеятельном состоянии, что встречается чрезвычайно редко в животном и растительном организмах. Кормление пен-тозанами на пентозурию не влияет. Пентозы входят в состав нуклеиновых к-т, и пентозу-рическая аномалия обмена имеет интерес скорее для области нуклеинового обмена, чем с точки зрения углеводного обмена; наибольшее значение имеют те количественные уклонения хим. превращений, к-рые происходят с углеводами при диабете. Нужно отметить, что при анализе как тканей, так и всевозможных жидкостей и выделений диабетика не удается найти каких-либо продуктов обмена, совершенно чуждых нормальному организму. Даже ацетоновые тела, представляющие столь резкую особенность тяжелого диабетического обмена, хотя и в значительно меньшем количестве, но все-таки встречаются и в нормальном обмене. Т. о. диабетический организм в обмене никаких новых особенностей по своему хим. строению веществ не создает, и все синтезы и разложения совершаются с образованием обычных промежуточных веществ. Это обстоятельство позволяет в нек-рой мере данные, найденные на здоровом организме, переносить и на диабетика. Сюда прежде всего относится то важное правило, согласно которому сахара при своих превращениях в обмене распадаются до образования трехуг-леродистых комплексов (молочная кислота, метилглиоксаль), распадающихся при дальнейшем ходе обмена до углекислоты и воды. С другой стороны, многие вопросы, существенные для нормального обмена, находят свое разрешение при рассмотрении диабетического расстройства. Такова напр. проблема образования сахара из тел неуглеводного строения. Если наблюдать влияние кормления глюкозой на выделение сахара у диабетика, то нетрудно убедиться, что под влиянием съеденной глюкозы больной выделяет больше сахара. Однако неправильно было бы думать, что всосавшаяся глюкоза не входит ни в какие дальнейшие реакции, а непосредственно выделяется мочой. Гликозурия увеличивается под влиянием не только глюкозы, но также и углеводов другого строения, так что приходится допускать по меньшей мере необходимость перегруппировок. Встает также вопрос о причине резкого повышения содержания сахара в крови, к-рое наступает не сразу после введения углевода в желудок, держится дольше, чем в норме, и говорит о расстройстве сложной регуляции гликемии. Левулеза (фруктоза) пищи хотя и увеличивает глюкозу мочи, но меньше, чем глюкоза. Зато от левулезы наступает более обильное отложение гликогена в печени диабетика. С другой стороны, и гликоген печени переходит в глюкозу мочи, доказательством чему служит то, что Клод-Бернаровский укол сопровождается положительным результатом только при печени, наполненной гликогеном. Т. о. вопрос о переходе углеводов пищи в глюкозу дает основание предполагать, что все углеводы пищи вероятно предварительно проходят стадий гликогена, к-рый диабетической печенью плохо фиксируется и легко отдается в виде глюкозы. Один из источников сахара т. о. найден в углеводах питания и гликогене печени. Что касается высоты концентрации сахара в крови, то объяснение ее в наст, время встречает большие трудности. Господствовавший прежде взгляд, что одного повышения концентрации достаточно для появления гликозурии, не может в наст, время поддерживаться полностью. Благодаря возможности химич. анализом проверять содержание сахара в капле крови удается часто наблюдать без всякой гликозурии большую гликемию в случаях сахарного мочеизнурения, при соотв. диете, при гипертонии и т. д. Гликозурия не находится в прямой связи с гипергликемией, встречаются даже случаи гликозурии, протекающие без повышения сахара крови (почечный и флоридзиновый диабет). Противоположная крайность — низкое стояние концентрации сахара в крови—наблюдается кроме случаев применения чрез^» мерно больших доз инсулина еще при Адди-соновой б-ни, фосфорном отравлении и экспериментальном вырезывании печени. Все экспериментальные наблюдения с определенностью указывают на зависимость высоты сахара крови от содержания гликогена в печени. Попытка найти изомеры сахара, из которых только один проходит через почку, осталась безрезультатной. В тяжелых случаях сахарного мочеизнурения выделение сахара не прекращается и тогда, когда б-ной долгое время питается только жирами и белками или даже просто голодает. Т. к. имеющиеся запасы гликогена быстро мобилизуются и теряются в виде мочевого сахара, то дальнейшее выделение сахара может поддерживаться только образованием сахара из других исходных продуктов. В виду этого приходится допустить возможность превращения в сахар белков или жиров или и тех и других. При этом безралично, происходит ли сахарообразование из этих веществ непосредственно или проходя предварительно стадий гликогена или другого вещества. Важно, что вещество неуглеводного строения может так или иначе перейти в углевод (понятно речь не идет об использовании готовой углеводной группы, заключающейся в нек-рых белках). Для решения этого вопроса может служить клин, наблюдение, состоящее в том, что чем больше белков тяжелый диабетик получает в пище, тем больше он выделяет сахара. К этому следует еще прибавить, что гликозурия нарастает также от кормления аминокислотами. Для образования такого экстрасахара пригодны аминокислоты с неразветвленной прямой цепью углеродов. Все эти вещества в обмене деза-минируются и затем, пройдя фазу окси- или кетокислот, используются как материал для образования сахара. Видную роль в этом процессе играет печень. Однако из всего этого еще не следует с безусловной необходимостью, что белок пищи переходит в сахар. Гликозурия может быть развивается не потому, что в обмен внедряется большое количество исходного материала, а потому, что или увеличивается общий обмен вследствие специфически динамического действия белка или от какого-либо иного влияния съеденных белков на ход обмена вообще. Все это тем более вероятно, что связь между экстрасахаром и пищевым белком выражается очень сложной, а не простой линейной зависимостью. Известная связь между количеством выделенного сахара и выделенного N при сахарной болезни имеется. Коеф. ^ (отношение глюкозы к общему азоту мочи) несколько меняется в зависимости от наблюдаемого случая и постановки опыта. Величина его то 2,8 то 3,65, иногда еще больше. Неправильно представление, что весь образовавшийся сахар переходит в мочу непосредственно и что реакция разложения белка быстро доходит до мочевины. Неудивительно поэтому, что, если вести расчеты, выходя из действительно наблюденных ^, то количество са- 72в хара, соответствующее 100 г белка, не больше 45—58 г. Без сомнения даже относительное постоянство ^ заслуживает большого внимания. Огромное количество работ, проведенных для разрешения проблемы о переходе белка в сахар, привело в наст, время к разрешению этого вопроса в положительном смысле. Коеф. ?- пользовались для своего доказательства также и сторонники взгляда, утверждающего, что сахар происходит из жира. По расчетам коеф. ^ при образовании углеводов из белка в лучшем случае равен 6,37 (Geelmuyden). Между тем различные наблюдатели опубликовывали цифры, значительно превосходящие предельное значение. Это значит, что разложившийся белок, давший N, не мог доставить всего выделенного сахара, к-рый мог в этих наблюдениях происходить только из жира. Но белковая молекула может быть и не распадается непосредственно, а проходит через ряд промежуточных ступеней, и кроме того время, потребное для образования и выделения сахара и мочевины, может быть различно. Кривые выделения белкового азота и белкового сахара оказались бы сдвинуты одна относительно другой. Другое доказательство в пользу образования сахара из жира со выводилось из дыхательного коефициента --~~ у диабетиков. При нормальном обмене наименьшая величина этого коеф. рассчитывается для жира и равна 0,707. Между тем в случаях тяжелого диабета весьма нередко можно встретить цифры, значительно более низкие. Это говорит за то, что кислород, связанный при дыхании (знаменатель коеф.), не выводит через легкие всего углерода окисленного жира (числитель коеф. RQ делается меньше). Это может быть в том случае, если жиры при окислении дадут углеводы, к-рые выделятся с мочой в виде глюкозы. Но кислород может быть связан и остается в организме, не давая С02, и при образовании ацетоновых тел или при другой фиксации 02 в организме. Обоим доказательствам образования сахара из жира, основанным на довольно сложных расчетах, противопоставляется обычно общеизвестное клин, наблюдение, что кормление жирами не увеличивает гликозурии. Это доказательство, основанное на чрезвычайно ярком клин, факте, оспаривается на том основании, что переход жира в углеводы может быть весьма медленным и кроме того жир организма мог бы в обмене давать другие продукты расщепления, чем жир пищи, и отличаться своим влиянием на гликозурию. Т. о. вопрос об образовании углеводов из жиров далеко еще не может считаться разрешенным. Он затрудняется прежде всего тем, что методика наблюдения за обменом жира разработана еще недостаточно. Представить себе переход жира в сахар трудно и химически, т. к. обмен жиров совершается через промежуточные вещества с числом углеродов, равным 4, а в обмене углеводов важную роль играют вещества с тремя атомами углерода. Продуктов промежуточного характера между этими двумя типами хим. веществ в обмене не обнаружено. Несмотря на это все же вполне доказано, что обратный путь — синтез жира из углеводов — в обмене веществ принадлежит к числу часто встречающихся.—Остается еще прибавить, что остаток глицерина, входящий в жиры, может повидимому являться в организме источником образования углеводов. Помимо вопроса об источниках сахара нужно еще решить, в чем заключается сущность диабетического расстройства. Даже трудно окисляемые вещества в организме диабетика разрушаются совершенно так же, как у нормального, и нет никаких оснований утверждать, что вообще окислительные процессы у сахарного больного понижены. Расстройство касается исключительно углеводного обмена. Имеются два допущения для объяснения сахарной болезни. Первое состоит в том, что сахар при диабете окисляется медленнее, чем в норме. Введение углеводов в кишечнике поэтому мало повышает RQ диабетика. Благодаря этому нормальный поток глюкозы из кишечника и тканей (главным образом печени) не успевает подвергнуться своевременному окислению, в результате чего наступает гликозурия. Но полной потери способности окислять сахар не наблюдается никогда. Это противоречило бы и современной точке зрения на сахар как на источник мускульной деятельности, которая у диабетика не угасает, а кроме того мышечные работы даже у тяжелого диабетика повышают RQ. Подтверждается замедление окисления сахара у ■диабетиков и наблюдениями над действием инсулина, под влиянием к-рого в эксперименте окисление углеводов в мышцах повышается. Второе предположение, выдвигав--мое для объяснения сахарного мочеизнурения, говорит об усиленном образовании сахара. Согласно ему углеводный поток, источники которого теряются в тканях и кишечнике, течет более бурно, чем в норме, заливает весь организм сахаром, который не успевает сгореть в соответствующих органах и потому удаляется через почки неиспользованным. Приспособления же, которые должны предупредить углеводное «наводнение» (гли-когенообразование), у диабетика не действуют. Такое предположение доказать труднее, чем замедление окисления глюкозы, хотя усиленная продукция сахара диабетиком, находящимся в трудоспособном состоянии, весьма вероятна. Her ничего невозможного и в том, что у сахарного б-ного поражено и сахарообразование и сахароис-пользование. Своеобразное уклонение углеводного обмена обнаружено в клетках опухолей. Всякая клетка для поддержания cbopix жизненных потребностей извлекает энергию из углеводов либо при помощи процессов окисления либо без помощи кислорода. В первом случае мы имеем перед собой дыхание, во втором то, что носит название брожения. Обе способности находятся друг с другом в сложном взаимоотношении. Если исследовать экспериментально обмен нормальных эпителиальных клеток, протекающий в присутствии кислорода, то легко доказать 7S27 72» их дыхание. Раковые клетки в этих же условиях кроме дыхания энергично перебра-живают глюкозу в молочную к-ту. Исследование тех же объектов в условиях анаэробиоза показывает, что и нормальный эпителий обладает способностью вызывать брожение, но в значительно меньшей степени, чем раковая клетка. Способность вызывать брожение в присутствии кислорода некоторыми авторами ставится в связь с атипичным ростом рака. Эта особенность считается характерной для ракового обмена, причем было доказано, что венозная кровь, оттекающая от раковых опухолей, богаче молочной к-той, чем нормальная. е. Фромгольд. Пат. анатомия нарушений углеводного обмена выражается лишь в отложениях в ткани гликогена, что бывает следствием общего расстройства углеводного обмена как при диабете (см. Диабет сахарный), так иногда при местных неправильностях этого обмена, напр. в опухолях и очагах воспаления (см. Гликогеновая инфильтрация). В случаях длительного нарушения углеводного обмена патолого-анатомическая картина может осложниться признаками общего нарушения питания в виде кахексии той или иной степени. ТП. Минеральный обмен. Минеральный обмен обнимает ряд явлений, касающихся всасывания минеральных элементов, их странствования по телу, их распределения по тканям и жидкостям организма и их выделения наружу. Валовое содержание (в процентах) минеральн. веществ в теле человека следующее (не считая 02):' Табл. 18. Элемент % Элемент o/ /o Са Na С1 Mg Ре 1,5 1,0 0,35 0,25 0,15 0,15 0,05 0,004 0,00004 Zn Si Al, Br, Cu, F, Mn As, B, Pb, Ti, Co, Ni 1 тысячные J ДОЛИ » десяти-V тысячные ) доли ^ стотысяч-i ные доли Не все элементы из указанного ряда относятся к «биоэлементам», нек-рые из них, как напр. Cu, Pb, Со и Ni, являются необязательной составной частью тела, по крайней мере у высших позвоночных. Минеральные элементы в теле представлены: 1) в виде отдельных элементов, входящих в состав органических соединений, как напр. S в цистине, Fe в гемоглобине, J в иодотирине; 2) в виде минеральных кислот и минеральных или органических солей, причем они (к-ты и соли) могут быть а) свободными и находиться в растворе в диссоциированном или недиссоциированном состоянии, б) связанными; эта связанность может быть либо слабой либо б. или м. прочной. Электролиты, минеральные катионы и анионы, при своем распределении по жидкостям и клеткам организма создают ту сложную систему концентрационных разностей, к-рая может определять собой тече- ние многообразных осмотических и диффузионных процессов в организме. Однако распределение отдельных электролитов в тканях, подчиняющееся принципу Доннана, обнаруживает, что их роль не ограничивается образованием отдельных осмотических напряжений; так, известно, что нормальная возбудимость мышц и нервов зависит также от определенного сочетания электролитов в ней (ср. состав жидкости Рингера, Локка и др.), причем изменения в отношениях между ними допустимы в очень узких пределах. Минеральный состав крови и нормальных тканей животного в общем характеризуется большим постоянством (при данных^ условиях), что зависит в основном от действия четырех факторов: от поступления неорганических соединений извне; от беспрерывного разрушения в тканях органических материалов в процессе катаболизма, освобождающих минеральные элементы в форме электролитов,—увеличивает концентрацию минеральных веществ в жидкостях организма; от постоянного выделения наружу минеральных веществ почками, кожей, кишечником и т. п.; наконец от накопления или отдачи минеральных элементов минеральными депо организма—костяком и вероятно многими другими тканями (в зависимости от концентрации в крови или под влиянием физ.-хим. свойств, например рН крови). Указанные четыре фактора представляют собой систему, где в каждый данный момент обнаруживаются взаимно противоположные действия: одни из них п о д н и м а-ю т уровень минеральных соединений в крови (напр. кишечник в периоды всасывания или депо-ткани в периоды обеднения крови минеральным содержимым), другие понижают этот уровень [напр. органы выделения (почки) или разные депо-ткани в периоды их пополнения минеральными соединениями]. При изменении питания организма минеральное равновесие сдвигается в ту или другую сторону лишь временно, до тех пор пока действие указанных факторов не восстановит его на прежнем уровне. При различных (длительно применяемых) системах питания i организмы могут показывать в своем минеральном составе соответствующие различия, особенно резкие в случаях дефективного питания. Способность организма создавать род подвижного равновесия между минеральными элементами — солями и ионами — крови и разных органов является в свою очередь результатом влияний на метаболизм отдельных органов, с одной стороны, и внешних воздействий—с другой. Напр. возникающая при работе мышц молочная к-та изменяет концентрацию водородных ионов, что в свою очередь меняет: соотношение между различными компонентами буферной системы крови (см. Буферные свойства); состав самой мышцы, лишая ее между прочим калия, к-рый выделяется из коллоидных связей молочной к-той (Burridge; 1906). Увеличение Н+ (ацидоз) в этом случае обусловливает также ббль-шую растворимость Са, в результате чего получается временная калькариурия. Из эндогенных влияний на минеральный обмен можно указать на отношения к нему со стороны эндокринной и нервной систем. Так, установлено, что после панкреатомии (у собак) развивается гипохлоремия, что ведет к уменьшению отдачи С1 мочой, содержание же его в органах меняется различным образом: оно возрастает в мышечной ткани и понижается в печени и коже (Meyer-Bisch, Ni; 1926, 1927), причем изменения в содержании хлора идут совершенно независимо от содержания Na. Кроме того в ряде опытов отмечалось, что адреналин понижает уровень С1 в лимфе (задержка С1 тканями!); обратно этому действуют препараты инсулина и «питугландоля». Об участии щитовидной железы в обмене Са и Р также имеется ряд указаний. По недавним работам Мизоками и Нишиму-ра (Mizokami, Nishimura; 1929) у собак имеет место усиление выделения Са после кормления щитовидной железой и уменьшение после тиреоидектомии. Аналогично этому и на людях установлено (Aub; 1929), что введение вещества щитовидной железы или тироксина вызывает увеличение выделения Са и Р. В связи с этим при гипертиреозных •состояниях (б-нь Базедова, аденома щитовидной железы) мы имеем повышение, а при гипотиреозах—понижение выделения Са и Р .{рисунок 5). Усиленное выделение Са и Р в описанных случаях происходит за счет костного вещества; в последнее время это подтверждается рентгеноскопически (при длительном базедовизме— большая прозрачность скелета, при микседеме— наоборот). Вместе с тем установлено также, что Са в крови понижается при базедовизме и у нормального человека ш Число случаен 2 15 ( Ыж и "Ряс. 5:I—болезнь Базедова; //—аденома щитовидной железы; /Л—норма; IV—микседема; V—пара-тиреопривная тетания. после приемов тироксина; обратное явление наблюдается при микседеме (Leicher; 1922). В обмене кальция и фосфора принимают также участие паращитовидные железы. Экспериментальное удаление их приводит к значительной гипокальцемии и высокой фосфотемии, что ставится в связь с .одновременно возникающими явлениями тетании. Однако следует заметить, что при тетании содержание извести в закончившем рост скелете, как и в мышечной ткани, не ниже нормы (Behrendt; 1926), так что дефект в обмене кальция касается не общего содержания этого элемента в организме, а его транспортирования по- телу. Одновременно выяснено, что при тетании выделение Са мочой понижается (ретенция Са); точно так же понижено и выделение фосфора, так что при наличии гиперфосфа-темии можно говорить о перегрузке тела фосфором и считать пониженную кальцемию следствием обогащения крови фосфатами [экспериментально показано, что введение фосфатов (человек, животные) значительно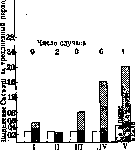
Рисунок 6: I—норма; //—послеоперационная тетания; III—норма, 50—5 5 единиц. гормона паращитовидной железы; /V—норма, 80— 100 единиц гормона пара-щитовидной железы; V — гинерпаратиреоидизм, Q, 41 г.
понижает кальций сыворотки (Binger, 1917; Gyorgy, 1924; a! Klercker, 1925; Gates, 1927)]. Совокупность всех этих явлений выражает алкалезное состояние организма, возникшее вследствие нарушения механизма регуляции при выпадении действия гормона паращитовидных желез. Наоборот, введение в организм этого гормона вызывает явления диаметрально противоположные (рис. 6). Связь минерального обмена с нервной системой составляет тему, еще очень мало разработанную., Влияние на минеральный обмен эндокринных органов предполагает эту связь через симпа-тич. и парасимпат. иннервацию.Непос-редственной иллюстрацией нервных влияний на минеральный обмен могут служить опыты Аль-перна (Alpern; 1925), показавшего изменение содержания К в слюне после раздражения симпат. нерва, а также исследования Нейш-лоса и Треллеса (Neuschloss, Trelles; 1924), к-рые обнаружили разницу в содержании К в мышцах, лишенных своей иннервации, по сравнению с мышцами с неповрежденными нервами. Далее по опытам ряда авторов перерезка п. splanchnici, удаление plex. coeliaci вызывают понижение Са в сыворотке без тетанических явлений, причем уровень Р остается без изменений; перерезка обоих nn. vagi повышает уровень Са. Всасываниеминеральных элементов. Минеральные составные части пиши подвергаются всасыванию гл. обр. в тонких кишках, причем из кишечного канала исчезают не только свободные соли, но и слабо связанные с органическими соединениями (напр. с аминокислотами). Процесс пищеварения содействует освобождению и более прочно связанных минеральных компонентов пищи. Для щелочных металлов и хлора отмечается очень быстрая и полная их всасываемость; у человека в нормальном стуле содержание С1 составляет 0,05 г pro die (Wendt). Иллюстрацией всасывания может служить опыт Гейден-гайна; изолированная двумя лигатурами петля пустой тонкой кишки собаки всасывала раствор NaCl след. образом (см. табл. 19). Очевидно из гипотонических растворов всасывание идет быстрее, чем в гипертонических; из гипотонических растворов, понятно, быстро всасывается осмотически вода, и раствор ■ концентрируется до содержания поваренной соли в плазме (около 0,65%), однако NaCl проникает в стенку кишки несмотря на то, что его концентрация в полости кишки меньше, чем в крови; причину этого явления надо искать в том, что диффузионные токи из полости кишки определяет не кровь Введено NaCl Содержание NaCll в растворе через | 15 мин. i Всосалось NaCl за 15 мин. см3 1 120 (120 % а /о СМ3 г % к введенному Гипотонич. 0,3 0,36 0,60 0,108 0,25 растворы 0,60 0,63 0,23 0,37 Гипертонич. \Ш 1,00 1,17 0,90 0,67 0,50 растворы J 120 1,46 1,75 1,20 1,31 0,44 своей концентрацией, а живой эпителий ворсинки, метаболические процессы к-рого нам пока неизвестны. Гипертонические растворы вызывают осмотически секрецию воды в полость кишки и угнетают всасывание соли. В наблюдаемых явлениях дело кроме того осложняется тем, что вероятно давление на содержимое кишки, возникающее благодаря работе мышц кишечной стенки, заставляет его продавливаться в межклеточные пространства кишечной стенки («фильтрация» жидкости), аналогично чему действует и своеобразный «насос» ворсинки. Переход С1 в кровь после всасывания при обычных пищевых режимах мало изменяет концентрацию этого элемента в крови—от 0,36% до 0,38%. Артериальная кровь богаче хлором, чем венозная кровь. Кратковременная гиперхлоремия (напр. 0,407% С1 в крови против 0,364% вначале) наблюдается только после дачи большой дозы NaCl. Общий же водно-солевой режим оказывает влияние на высоту С1 в крови; длительное низкое содержание поваренной соли в пище снижает и в крови содержание С1 до нижних пределов его нормальных колебаний. Постоянное потребление очень соленой пищи повышает содержание С1 в крови. В первые часы пищеварения наблюдается нек-рое понижение С1 в крови под влиянием увеличенной секреции желудочного сока. Подобно хлору натрий и калий после всасывания в зависимости от содержания их в пище, скорости всасывания и водного режима могут увеличивать свою концентрацию в крови на нек-рое время (нормальные колебания Na и К в плазме идут в границах 0,28—0,35%, resp. 0,016—0,024% у человека и близко к этим числам у др. млекопитающих), причем заметен ясно выраженный антагонизм между К и Na: увеличение содержания одного из них в крови уменьшает содержание другого. Выделение хлора и щелочных металлов из крови происходит гл. обр. мочой, а также путем потоотделения; при значительных потениях организм теряет не малое количество хлоридов; по Тютеру (Tuteur), до 4% общей потерихлора идет через кожу.При очень обильном потоотделении вследствие механической работы при высокой t° (напр. при восхождении в гору в жаркое время) по данным Конгейма, Тоблера и др. (Cohnheim, Tobler) пбтом терялось до 10 г NaCl в течение немногих часов. По данным Aggazzotti при путешествии в горах потеря С1 через кожу была так велика, что в течение нескольких дней NaCl в моче вовсе не появлялся и наблюдалась значительная гипохло-ремия (до0,43% NaCl вкрови, resp. 0,17 %С1). табл. 19. Вопрос об усвоении каль-ция и фосфора затруднен, т. к. оба эти элемента выделяются из организма не только почками, но и кишками, причем последним путем удаляются нередко очень значительные количества этих элементов; поэтому при обычных исследованиях усвоения Са и Р, когда производится сравнение содержания их в стуле и в пище, вовсе нельзя решить, какая часть происходит из невсосанных солей пищи и какая часть образовалась в результате выделительной функции кишечника. Из экспериментов, доказывающих способность кишки всасывать Са и Р, кроме старых опытов (Фойт, Форстер, Raudnitz; 18"<5—1893) можно указать также на опыты Аристовского,Уол-ша, Айви (Walsh, Ivy; 1925—1928) с фистульными собаками; оказалось, что ре-зорпция Са и Р происходит в ileum, а для Р еще и в jejunum, но 40—50% введенных элементов покидает тонкие кишки, не всосавшись. Табл. 20. Пища: 600 см» молока В химусе через фистулу выделилось Процент выделенного к введенному Са Са Са % 0,595 0,595 0,543 0,578 0,83 0,83 0,84 1 0,84 | 0,224 0,21!) 0.244 0,230 0,308 0,458 0,493 0,412 37 36 45 39 37 55 58 47 600 i мяса | 1,34 1 1,34 0,083 | 0,083 ! 0,092 | 0,089 0,187 0,159 7 7 224 190 Из таблицы 20 видно, что при бедной кальцием пище получается очень значительная потеря этого элемента через фистулу, что иллюстрирует экскреторную способность кишки. Часто наблюдалось и у человека значительное превышение Са в кале над количеством его в пище. В опытах Ауба (Aub) в среднем за 3-дневный промежуток каловый Са (у 13 индивидуумов) составлял 0,60 г против 0,33 з Са в пище. Подобные же наблюдения обнаруживают экскреторную функцию кишок и по отношению к Р (таблица 21). Табл. 21. Авторы Tlgerstedt Р пищи . | 0,027 I | 0,120 Renvall.....<\ 0,112 I I 0,072—0,096 Zuntz...... I 0 Р кала 0,134 ►0,223 0,229 0,065—0,044 В опытах Р. Берга (1911) повышенное выделение Са и Р кишкой длится нек-рое время после прекращения притока Са и Р в пище. Всосавшийся через кишечник Са не дает заметных изменений уровня Са в кро- ви; только очень большие дозы Са (до 30 г безводного СаС12) вызывают легкий подъем его, так что содержание Са в плазме крови является очень устойчивым (9—13мг%почти у всех млекопитающих), что указывает на деятельную работу кальциевых депо (кости), быстро реагирующих на изменения кальце-мии. Что касается фосфора, то прием его в пище вызывает, хотя впрочем не всегда, повышение Р крови;у кроликов и маленьких детей прослежена после дачи фосфатов постепенно увеличивающаяся фосфатемия, достигающая через 2—3 часа максимума (до 180% против первоначального). Колебания свободных фосфатов в крови претерпевают очень значительные вариации, по разным авторам от 4 до 18 мг%; влияние оказывает и общее питание; при бедной фосфором пище неорганические фосфаты крови понижают свой уровень. Подобно другим элементам всасывание железа происходит в тонких кишках; это доказано разными методами, между прочим морфологически (микрохим. реакция на железо). (Относительно вопроса о формах,-в виде к-рых всасывается железо, см. Железо.) Как и у предыдущих элементов, очень зат-"рудняет решение количественной задачи об усвояемости железа тот же факт, а именно, тальной эритропении наблюдалось достижение нормы путем усиленного введения железа с пищей (см. ниже патологию обмена;, что говорит за влияние пищевого железа на эритропоэз. При нормальном состоянии крови усиленный подвоз пищи, богатой железом, может увеличить содержание этого элемента в его депо - тканях (костный мозг, селезенка, печень), не отражаясь на содержании Fe в крови; так, при подкармливании мышей железом содержание последнего в их теле возрастало в сравнении с контрольными животными в 3 раза. Всосавшиеся и перешедшие в кровь минеральные вещества распределяются по телу, переходят в соответствующие депо-ткани, увеличивая их массу, заменяют возникшие пробелы в других тканях, не нашедшие же себе применения неорганические материалы выходят из организма указанными выше путями. Различные ткани предъявляют организму неодинаковые требования на минеральные компоненты, характеризуясь б. или м. постоянным составом. Наиболее постоянен он в костях, хотя и здесь разпые исследования дают неоднородные числа и даже у одного и того же животного симметричные кости обнаруживают неодинаковое содержание минеральных элементов. Ткани Кость.........j 1780 Мышцы поперечнопо- | лосатые........ 80 Мышцы гладкие . . . i 112 Мозг..........100—180 белое вещество . . . . ! — серое вещество . . . . I — Печень.........! 60 Кожа (к сух. вещ.) . ! 100—143 Селезенка.......| — Кровь (см. Кровь) \ что кишка является одновременно и всасывающим и экскреторным органом для железа. Принимается, что желудок и тонкие кишки производят резорпцию, толстая кишка— экскрецию железа, однако твердых оснований для такого распределения ролей в кишечнике в сущности не имеется. Кроме кишечного эпителия Fe выделяется также желчью, у человека до 1 мг pro die; эта величина считается постоянной, не зависящей от притока железа в пище или парентерально, т. к. она происходит от распада эритроцитов; возможно, что это железо в тонких кишках частично вновь всасывается. По исчислениям Гофмейстера (Hofmeister) от распада эритроцитов должно освобождаться 40—50 мг Fe, из к-рого мочой выделяется 1—2 мг, остальное—через кишечник. Всосавшееся в кишке Fe проникает как в кровь, так и в лимфу; это прослежено морфологически (микрохим. реакции), однако не было возможности до сих пор заметить обогащение этих жидкостей железом даже при условии максимального всасывания; дело в том, что плазма крови и лимфы считается практически свободной от железа (по Erben'y 1 кг кровяной плазмы содержит 23 мг Fe; 1 кг белых телец—95— 122 мг) и в обычных пробах крови прибавка всосавшегося железа не поддается учету. Что же касается количества Fe в цельной крови, то оно зависит от содержания Fe в эритроцитах (см. Кровь) и от количества последних в крови. Влияние различных пищевых режимов на содержание Fe в крови проследить трудно, только в редких случаях экспериментальной^ анемии и эксперимен- Т а б л. 22. Содержание минеральных элементов в некоторых тканях (по разным источникам) в мг %. Na Са ! Мё 320 250 240 400 58—119 7 940 7,5 5 2—17 26—78 5—15 12—21 8—14 9—17 7—10 Fe 15 ! 2 ! 5—10: С1 70 120 150 3—601100—200 — '240—480 72 j 161 р 3 850 — (224) (9) (210) (92) Кость характеризуется большим постоянством своего состава, что в свое время вызывало попытку установить определенную хим. «формулу» костного вещества (Норре-Seyler). Однако по взгляду Гофмейстера (1911) наблюдаемое постоянство состава кости зависит от постоянства состава питающей ее среды—кровяной жидкости. Неправильное питание организма, разные виды солевого голодания, могут изменить содержание минеральн. соединений кости (см. ниже патологию). Изменение рН крови в кислую сторону вызывает растворение минеральных веществ кости и усиленное выделение Са и Р через мочу, что характерно для ацидоза. «Искусственно» вызванный ацидоз (дача молочной к-ты, щавелевой, слабой серной и т.д.)ведет к обеднению костей известью. Из таблицы ясно видно, что кость очень богата минеральными элементами и представляет собой действительно депо-ткань для этих элементов, особенно для Са и Р. При голодании костная ткань сильно «тает», лишаясь своих минеральных элементов (уменьшение костной золы) (Morel, Mouii-quand и др.; 1921—1925) и в частности Са и Р (Iwabuchi; 1922). Длительный недоста- ток в пище кальция может привести к полному рассасыванию нек-рых костей (Лавров, Ярусова). Конечно кроме костной массы минеральные элементы освобождаются и из других тканей, разрушающихся в порядке катаболизма. Руководствуясь потерей белка в теле (по азоту) и считая, что этот белок происходит от разрушения мышечной массы, можно подсчитать, сколько должно бы освободиться при этом минеральных элементов (зная минеральный состав мышц) и сравнить их количества с таковыми мочи и кала подопытного лица. В таблице (наблюдение Gamble над голодавшей девочкой) каждый минеральный элемент выражен в децимиллиэквивалентах, другими словами, эквивалентным количеством см3 п/10 раствора. Табл. Периоды голода (по 3 дня) Na Mg о,™ £ « о о i° я io о S3 if! о м си п о В ев So й- аю art О) '<*•£ Я CD Е- >s) Мм К <D Од * Ив Яо да и 13,3 11,5 10,3 0,3 0,9 1,1 0,7 1,0 1,0 « 0,6 1Д 0,9 ]_ Из таблицы 23 видно, что для К и Mg отношения найденного к вычисленному очень близки к 1; это указывает на то, что разрушаемая при голодании мышечная масса действительно является источником этих элементов, между тем как для Са найденное число превышает вычисленное более чем в 10 раз; следовательно источником Са в этом случае должна быть другая ткань; таковой тканью является кость. Сравнивая минеральный состав других тканей, мы видим, что большинство из них характеризуется сравнительно большим содержанием калия и бедностью кальцием; по содержанию железа первые места занимают—уступая все же значительно кости—печень и селезенка, к-рые в отношении этого элемента являются депо-тканями значительные колебания Fe в печени зависят от «доставки» элемента этому органу, причем эта «доставка» может быть экзогенная (из пищи) или эндогенная (из других органов).— Привлекала внимание исследователей и кожа в виду больших колебаний в ее минеральном составе. Кожа позволила обнаружить в отношении щелочных металлов и хлора нек-рую зависимость от рода пищи. Нек-рые авторы рассматривают поэтому кожу как депо-ткань для только-что названных элементов. Кроме указанных в таблице элементов в разных тканях были находимы более редкие элементы, появление которых носит повидимому случайный характер, напр. Zn (3—5 мг%), Си и А1 в мышцах, Zn до 15 мг% (человек) и 34 ли% (лошадь) в печени, там же Си до 12 мг%, Sn 10 л<г%,иМп, As, В, F и Др. вдоляхл*г%. Значение минеральных элементов в питании не ограничивается только абсолютным уровнем того или другого элемента в теле или пище, но также отношением этих эле- ментов друг к другу. Так, Л. Рединой отмечено оптимальное отношение Na : К, как 5:1 (на белых мышах). Выяснено также, что Са оказывает благоприятное действие на использование Fe в организме: при увеличении Са в пище можно сохранить равновесие железа на более низких уровнях. К вопросу же о взаимных влияниях минеральных элементов относятся случаи антагонистических явлений. Так, при хорошо сбалансированных диетах дача магния обусловливает потерю Са в организме; неблагоприятный эффект от инъекции Mg пропадал после последующей инъекции Са. Са вообще может коррегировать нарушение неорганического равновесия; при всяких аномальных эффектах, вызванных экспериментально щелочами (Na, К, Mg), Са действует восстанавливаю-23 щим (равновесие) образом. Сюда же относится классический опыт Бунге (Bunge), вызвавшего обильное выделение Na из организма дачей большого количества К. На основании балансовых наблюдений над беременными женщинами и на основании данг ных анализов человеческих плодов в разные сроки развития можно было составить следующие диаграммы для Р и Са (рис.7 и 8). Отношение минеральных кислот к минеральным основаниям в кровяной жидкости и в организме вообще. Сопоставляя анализы разных исследователей, дающих минеральный состав человеческой плазмы или сыворотки, нек-рые авторы (Heubner, Kramer, Marrack и др.) могли сравнить кислотные и щелочные эквиваленты кровяной жидкости (плазмы). Разница между к-тами и основа- са К са О Н 13,4 10,4 11,3 грСа ао д t/, s^y & Jk у У ( См fXH'. -^eY( Ж? '<?' / ^ Tlcii Wb3« £ав^ C"""S Ыв& Неде 17 ли бе 2< реме ) иност 24 И р 28 ИС. S2 8« ниями, представленными в милли-эквива-лентах, для плазмы составляет 151—135 = =16 милли-эквив. (по другим авторам от 10 до 25) в пользу оснований, причем к-ты (135 милли-эквив.) полностью покрываются натрием (139 милли-эквив.). Избыток щелочей (16 милли-эквив.) связан вероятно с кислыми белками, м. б. также с высшими жирными к-тами. Такая установка в организме, близкая к нейтральности [рН=7,36—7,32 (37°)], очень существенна для ряда физиол, процессов промежуточного обмена (см. Алкалоз и Ацидоз). Все случаи, нарушающие эту установку, требуют соответствующей реакции со стороны организма; в первую очередь используется буферная система, затем работа дыхательного центра, побуждающая учащенным дыханием эвакуацию С02, в дальнейшем происходит растворение запас-
ных щелочей из тканей и наконец —отщепление аммиака от аминокислот (resp. белков) в качество нейтрализующего основания; -в этой последней реакции существенную роль играет печень, а во всех стадиях развития ацидоза также и почки, поскольку они способны выделять кислую мочу. A priori можно было бы представить, что пища, содержащая в себе разные отношения минеральных к-т к основаниям, способна произвести нек-рые сдвиги нормы рН в ту или другую сторону, и только компенсаторной работой депо и органов выделения норма эта может сохраниться. С этой точки зрения для организма вовсе не безразлично, каково отношение к-т к основаниям в пище. Вопрос этот давно начал изучаться в Америке в лаборатории Генри Шермана (1911—12), причем в первую очередь исследования касались обычных пищевых средств для установления их «кислотности» и «щелочности». Для иллюстрации приводится краткая таблица из работ Шермана.
Продукты Перевес кислотных элементов в см* п/10 кислоты на 100 г продукта Продукты Кукуруза . . . Сухари ..... Рыоа...... Яйца...... Яичный белок Яичный желток Мясо ...... Ры5а...... Мука овсяная . Рис....... Пшеница цельная Пшеничная мука Яблоки ..... 6,95 7,81 9,89 11,10 5,24 26,69 0,05—13,91 9,5—16,0 12,93 7,8 ! 9,66 ! 11,6 ! Бобы....... Свекла ...... Капуста..... Цветная капуста Молоко..... Апельсины . . . Горох ...... Картофель. . . . Сливы ...... Редиска ..... Изюм....... Турнепс ..... Из таблицы 24 видно, что к кислым продуктам относятся мясо, яйца, злаки, к щелочным—молоко, плоды, овощи. Т. о. была введена новая сторона в оценке пищи. Требовалось выяснить, каков должен быть оптимум отношения к-т к основаниям пищи. Однако до сих пор ни физиология, ни клиника, ни зоотехния не дали еще определен- ных разработок в этом вопросе. С другой стороны, вопрос о потребности человека в том или другом минеральном элементе давно разбирался в физиологии, причем в основе всех исследований лежало предположение, что для человека данного возраста должен быть один какой-то минимум и оптимум для каждого минерального элемента. Такая постановка вопроса по существу своему неправильна, т. к. кроме отношений к-т к основаниям в пище требуется изучить еще сложный клубок зависимостей потребности минеральных компонентов пищи от ее органических компонентов, от водного хозяйства организма, от отношений щелочных элементов друг к другу, от компенсаторных способностей человеческ. организма по отношению к установке равновесий в системе: «депо-ткань—кровь—потребляющий и выделительный орган», а эти способности в свою очередь связаны с проф. привычками, упражнениями и вообще со всеми условиями жизни. Поэтому всякие указания авторов на уровень потребностей человека в отдельных минеральных элементах надо считать провизорным решением вопроса. Так, Шерман поставил 97 опытов на людях с целью определить, при каком уровне кальция (также и по отношению к другим элементам) можно было получить равновесие [т. е. когда баланс Са (Са пищи— Са выделений)=0]. При этом выяснилось, что у разных субъектов минимальный уровень Са, годный для установка равновесия, чрезвычайно различен—от 0,27 до 0,82 г на человека в день. Была взята средняя величина: 0,45 з. Подобные опыты ставились и для фосфора, где среднее оказалось 0,88 г на человека в день (колебания 0,52—1,20), и небольшое количество опытов для железа, давших колебания от 6 до 16 мг на человека в день; здесь уже остановились на округленном числе—15 мг. Конечно Шерман и для фосфора и кальция предложил увеличенные значения, а именно для Са—-0,68, для Р— 1,32 г на человека в день. Эти величины приняты в Америке Как ГИГ. НОРМЫ. Б. Лавров. Патология минерального <й5-меяа. Минеральные соли lbs-ляются составной частью тканей и жидкостей; эта их пластическая роль выявляется при голодании, когда вместе с выделением продуктов распада органических составных частей организма выделяются и минеральные соли. Лишение минеральных солей ведет к гибели животного. Длительное преобладание в пище одних солей и отсутствие других пови-димому до известной степени | | может изменить минеральный —:--------— состав организма (деминерализация Ноордена). Нек-рые авторы считают.что известное соотношение минеральных солей в пище с преобладанием кислых лли основных валентностей может влиять шя. общий обмен веществ в организме (Bfflcg), на течение болезненных процессов, заживление ран (Sauerbruch, Hermannsdwfer). Минеральные соли имеют регуляторяое значение Табл. 24. Перевес щ лочных элементов в еле8 п/к> кислоты на юо г продукта 23,8 10,9 4,3 5,3 2,37 6,61 7,0 5,5—7,2 24,4 2,9 23,7 27,7 для поддержания постоянства реакции тканей и жидкостей организма (см. Буферные свойства, Резервная щелочность). Благодаря буферам крови и тканей скопление в организме кислот или оснований не ведет к изменению активной реакции крови (компенсированный ацидоз, компенсированный алкал ез). Сдвиг активной реакции в кислую или щелочную сторону бывает крайне редко, б. ч. незадолго до гибели организма. Но при компенсированном ацидозе несмотря на но-измененное рН крови происходит изменение состава крови: понижение резервной щелочности, уменьшение напряжения углекислоты, понижение кривой связывания С02, кроме того в крови уменьшается количество оснований, преимущественно Na. Иногда при диабетическом ацидозе такие потери Na делают невозможным борьбу против ацидоза только при помощи инсулина, но требуют введения NaHC03. Обратный процесс происходит при алкалезе. Минеральные соли регулируют осмотическое состояние (см. Осмотическое давление) тканевых жидкостей, особенно крови. Понижение точки замерзания крови с А=—0,56° на несколько сотых градуса указывает нередко на значительные нарушения в организме. Преходящее повышение осмотического давления наблюдается при больших потерях жидкости (потение при работе в горячих цехах, при холере), длительное—гл. обр. при недостаточности почек. Концентрация минеральных солей (электролитов) имеет большое значение для стабилизации коллоидов, имеет влияние на состояние возбудимости клеток. Благодаря исследованиям Рингера и Леба (Ringer, J. Loeb) установлено, что правильные функции клеток поддерживаются смесью ионов К, Na, Са, Mg, находящихся в определенных соотношениях: изменение этих отношений может вести к возбуждению или к параличу. Изменения этих отношений тесно связаны с заболеванием желез внутренней секреции и с нарушением деятельности нервной системы. Краус и Цондек (Kraus, Zondek) связывают действие К с блуждающим нервом, действие Са—с симпат. иннервацией. Понижение Са и повышение К в крови наблюдается при тетании как паратиреоприв-ной, так и идиопатической и гуанидиновой; изменение электролитов сопровождается одновременно алкалезом; уменьшение Са встречается при гипертиреозе, увеличение—при гипотиреозе. Изменения в составе минеральных солей крови имеют место при аллергических состояниях, инфекциях, лихорадке. Часто имеет значение не абсолютное содержание того или другого иона, а взаимоотношение двух ионов, например -^ при артериосклерозе (см. Кровь). Специальный обмен различных минеральных солей и ионов. Хлористый натрий. Задержка хлористого натрия и выделение его тесно связаны с водным обменом (см. ниже) (см. Отек, Нефрит). При крупозной пневмонии и многих лихорадочных процессах бывает значительная задержка NaCl: моча почти не содержит поваренной соли; одно- временно в крови количество NaCl не только не повышается, но, наоборот, содержание его понижается (Monakov).—Кальций. При голодании количество Са, выделяемое мочой, увеличивается, выделяемое калом-— уменьшается. Параллельно повышению выделения Са уменьшается выделение Mg. Мнение о деминерализации, потере Са при tbc неподтвердилось. Обмен кальция тесно связан с обменом фосфора: кости являются главным местом отложения фосфорнокислого кальция. Кроме фосфорнокислого кальция в костях содержится и углекислый кальций, к-рый может извлекаться из костей при большом количестве к-т в организме для насыщения их (напр. при диабете). При пат. процессах, связанных с костной системой и характеризующихся недостаточным окостенением или расплавлением костной ткани, как при рахите и остеомаляции,. количество Са и Р в костях уменьшается, но соотношение Са++ и Р04'" не изменяется. В флоридном периоде рахита выделение Са и Р повышено (Са и Р в крови, см. Кровь). Одновременно с повышенным выделением Са наблюдается задержка Mg. При остеомаляции выделение Са и Р не представляется однородным; в костях наблюдается, как и при рахите, уменьшение Са и Р, но соотношение их не меняется (см. Кровь). При различных формах объизвествления и окостенения (myositis ossificans, calcinosis и др.) взаимоотношение между Са и Р и CO., такое же, как в нормальных костях. Отложение известковых солей повидимому является вторичным процессом; первично же ^развивается регрессивное изменение тканей. Часто встречается отложение известковых солей в различных участках организма, поврежденных инфекцией или токсическим воздействием (туб. поражение, тромбы, артерио-склеротические сосуды). Даваемый за последнее время в чрезмерном количестве Vigantol (витамин D) часто ведет к объизвествлению сосудов, что может зависеть от того, что ви'гантоль изменяет соединительную тканьг в к-рой затем происходит отложение известковых солей. Фосфор. При голодании или недостаточном питании количество фосфатов в моче увеличивается; путем вычисления соотношения выделяемых N:P выяснилось, что фосфаты мочи не только органического, но и неорганического происхождения. Большие потери фосфора встречаются при кормлении препаратами щитовидной железы и при болезни Базедова. Задержка фосфатов наблюдается наряду с задержкой хлоридов при острых лихорадочных заболеваниях, при нефритах (см. Кровь). Фосфатурия не является б-ныо обмена в узком смысле; она, как и оксалурия (см. ниже), относится к аномалиям формы их выделения. Фосфатурия— выделение мочи, в к-рой фосфаты содержатся в виде осадка. Осадок б.ч. аморфный [средний фосфорнокислый кальций, Са;1(Р04)2^ углекислый кальций], отчасти кристаллический [СаНР04, двуметальный фосфорнокислый кальций; трипельфосфат—Mg(NH4). .Р041. Моча б. ч. имеет щелочную реакцию. Между образованием осадка и общим количеством фосфатов мочи нет никакой прямой зависимости. По Зендтнеру, Тоблеру (Send-tner, Tobler), при фосфатурии количество Са в моче повышено, количество фосфатов нормально. Повидимому выпадение фосфатов зависит от состояния коллоидов мочи (Lichtwitz). Некоторые авторы (Minkowski, Лихтвиц) рассматривают фосфатурию как секреционный невроз, выражающийся в утрате способности почек выделять кислую мочу.—О к с а л у р и я—появление в све-жевыпущенной моче кристаллов щавелевокислого кальция. Повышенное выпадение в осадке оксалатов не зависит от концентрации щавелевокислых солей; выпадение щавелевокислого кальция может иметь место в кислой, щелочной и нейтральной моче. Клемперер и Фишлер (Klemperer, Fischler) ставят растворимость оксалатов в зависимость от содержаниямагнезиальныхсолей,но магнезиальные соли не увеличивают растворимости оксалатов,хотя и могут доизвестной степени подавить оксалурию. Лихтвиц считает, что оксалурия, как и фосфатурия, зависит ОТ СОСТОЯНИЯ КОЛЛОИДОВ МОЧИ. Ю. Гефтер. П а т. - а н а т. проявление нарушений минерального обмена может быть двояким: или оно представляет собой результат уменьшения количества солей в ткани или оно выражается в отложении, выпадении солей в какую-нибудь ткань или в полость.. Первое касается кальция, уменьшение количества к-рого в костной ткани имеет место при ряде специальных «малятических» б-ней костной системы, дающих своеобразные пат.-анат. картины (см. Остеомаляция, Остит фиброзный, Рааит). Выпадение и отложение солей может относиться также к кальцию (см. Известковые отложения, метастазы), а также и к другим солям (см. Подагра, Конкременты). Пат. анатомия нарушений сбмена железа — см. Анемия, Гемо-сидерин, Гемохроматоз, Железо—железа отложение. VІІІ. Водный обмен. О. в. водный в сущности является основным видом промежуточного обмена, т. к. вода есть главнейшая составная часть организма. У взрослых вода составляет 58—64% общего веса их тела, у новорожденных детей достигает даже величины в 74% и более. Наряду с этим почти все хим. процессы, разыгрывающиеся в О. в.,—диффузия растворимых веществ, влияние осмотических сил, набухание и отбухание белков крови и тканей, образование сеьретов и экскретов—■ протекают либо в водных растворах либо при непременном участии воды. В отдельных органах и тканях отмечается чрезвычайная устойчивость процентного содержания воды. Из 50 примерно веществ, входящих в состав мечи и крови, лишь единичные могут быть растворены также в др. растворителях кроме воды (Гендерсон). Кроме того молекула воды благодаря своей диполярно-сти ярляется прекрасным диэлектриком (см.), благодаря чему создаются условия для наилучшего растворения в ней электролитов и их диссоциации (см. Диссоциация электролитическая). Вода обладает также максимальным (после ртути) поверхностным натяжением, чем объясняется многообразие явлений адсорпции из водных растворов. Наконец диполярность водной молекулы и непрочность ее соединений обусловливают собой ее диспергирующее влияние на коллоидные взвеси (см. Гели). Максимальное содержание воды отмечается в почках (82,7%), в мышцах (75—80%) и нервах (до 82%), а . минимальное—в жировой ткани (до 29%) и в зубной эмали. В абсолютных величинах в мышцах содержится около половины всей воды, входящей в состав организма (табл. 25), причем до наст, времени остается неясным, являются ли мышцы местом скопления водных запасов организма, его депо, либо же, что более вероятно, вода служит постоянным участником весьма интенсивных биохим. процессов, происходящих в мышцах. Содержание воды в органах поддерживается с исключительным постоянством, т. к. искусственное отнятие или потеря 10% общего количества воды ведет к гибели организмов. Табл. 25. Процентное содержание воды в главных органах человека и распределение воды в них. Органы и ткани Кровь..... Жир...... Коша...... Костная ткань Мышцы . . . . Нервная ткань Кишки..... Сердце ..... Печень ..... Легкое ..... Селезенка . . . Почки..... Прочие..... Количе- Процент ство веса воды в % тела Процентное отношение к общему колич. воды 77,9—83,0 29,9 31,9—73,9 22—34 73,0—75,7 75—82 73,3—77,0 79,2—80,2 68,3—79,8 78—79 75,8—86,0 77—80 4,9 23 6,2 16 37 2,7 4,7- 6,6- 9,0- 47,7-2 3 2 1 2 0 0 11 - 9,0 ,3 -11,0 -12,5 50,8 .7 ,2 ,5 ,8 ,4 ,4 ,6 ,0 В промежуточном обмене участие воды возможно либо в качестве растворителя молекулярных частиц либо как гидратаци-онной воды, входящей в состав протеинов плазмы, клеточной протоплазмы, лимфы и межклеточной субстанции как гидрофильных коллоидов (см. Гели). Вода, играющая роль растворителя, отличается большой подвижностью, «текучестью» и легко воспринимается и отдается тканями («межклеточная» вода); гидратационная же вода, в особенности в клеточной протоплазме («конституционная» вода), является основной составной частью тканевых коллоидов, и потеря ее подчас равносильна гибели данной ткани. Вместе с тем несомненно, что воды, «свободно» циркулирующей в организме, не имеется, так как даже самые жидкие пат. трансудаты являются коллоидными растворами, содержащими еще и некоторое количество солей.— По своему происхождению участвующая в О. вода может быть различной природы. Ббльшая часть ее всасывается из кишечника и поступает в кровь, лимфу и ткани—т. н. пищева -вода. Далее, в процессе обмена и в перв^ очередь при окислительных процессах пример окисление углеводов, жирных в самом организме образуется т. н. от пионная вода; также образуется вода в результате реакций нейтрализации. Наконец тканевые и кровяные протеины способны под влиянием гидростатических и электроосмотических сил отбухать (Entquellung немецких авторов), отдавая свою конституционную воду. В наст, время еще далеко не выяснены все направляющие силы и пути, по к-рым осуществляется промежуточный обмен воды. Так, принятая внутрь вода не всасывается в желудке, однако ее поступления в желудок уже достаточно для того, чтобы вызвать вазомоторную кожную реакцию и усиление кожной перспирации. С переходом той же воды в кишечник начинается ее всасывание, быстрота к-рого определяется ионным и молекулярным составом выпитой жидкости. Далее всосавшаяся из кишечника пищевая вода не попадает тотчас же в общий ток крови и лимфы, а задерживается частично в печени, причем степень этой задержки зависит как от характера раствора, в составе к-рого вода всасывается (вода per se, солевой или сахарный, гипер- или гипотонический растворы), так и от состояния паренхимы печени и ее кровеносной системы. С физиол. стороны остается неясным, обусловлена ли подобная «шлюзовая» роль печени для воды специфическим устройством ее венозной системы (легкая проницаемость ее капиляров, наличие жома или венозных заслонок в месте выхода из печени печоночных вен) или же тому причиной исключительная рыхлость паренхимы печени, способность ее клеточных элементов к набуханию и наконец наличие огромного количества тканевых щелей в печени. Значительная часть всосавшейся из кишечника воды, в особенности после обильного питья, пройдя печень, поступает в круг кровообращения. Однако незначительность и кратковременность гид-ремии (см.) даже после обильных водных нагрузок (1—lVs л) свидетельствуют о том, что почти вся резорбированная вода быстро удаляется из крови. При помощи взвешивания и учета диуреза можно легко убедиться в том, что главная масса выпитой воды переходит в ткани и в лимфу и затем лишь постепенно отчасти выводится с мочой.Заслуживает внимания,что чистая вода легче удаляется из крови, resp. обусловливает меньшую гид-ремию, нежели изотонический раствор поваренной соли. Дальнейшее передвижение воды через сосудистую стенку и далее через полупроницаемые клеточные оболочки осуществляется в основном тремя силами: гидростатическим давлением, существующим в сосудистом русле, осмотическим давлением по обе стороны капилярной стенки и онкотическим давлением коллоидов крови и тканевой жидкости. Совершенно очевидно, что все попытки объяснить выхождение воды из кровеносной системы одной лишь фильтрацией ее под давлением (Ludwig) или влиянием осмотических сил (Starling) оказались тщетными. Мало того, следует учесть, что упомянутые три направляющие силы водного обмена имеют своей точкой приложения капилярный эндотелий, resp. клеточные мембраны, безусловно не подчиняющиеся полностью всем тем закономерностям, к-рые были получены при модельных опытах in vitro (Loeb). В то время как лимфа, являющаяся фильтратом кровяной плазмы, содержит соответствующее составу крови количество молекулярных частиц, происходящие из той же кровяной плазмы спинномозговая жидкость, серозная жидкость брюшины, а в патологических условиях и отечная жидкость, имеют весьма отличный от кровяной плазмы количественный солевой состав. Объяснения этим отклонениям даетфиз.-хим. биология,к-рая учит,что активная реакция среды по обе стороны мембраны, качественный и количественный состав электролитов определяют собой проницаемость упомянутых «барьеров» (гемато-энцефали-ческий, брюшинный, плевральный и пр.) для ряда ионов и неэлектролитов, а с ними и для их растворителя — воды. Наряду с этим небезынтересно также отметить, что в большинстве органов и систем непосредственный обмен воды между кровью и паренхиматозными клетками вообще невозможен, так как везде между ними имеется большее или меньшее количество соединительной ткани. Последней принадлежит роль не одной лишь опорной ткани: коллоиды ее элементов обладают исключительной способностью к адсорпции кислых валентностей (гистиохимически они ацидо-фильны), последствием чего является быстрый приток жидкости с положительно заряженными Н-ионами в межклеточное вещество и его набухание. Больше того, новейшими исследованиями (Hulse, Hueck) подвергнуто сомнению само наличие тканевой жидкости в межклеточных пространствах. С этой точки зрения коллоиды межклеточного вещества (припомощи т. н. «тканевого тока») сами осуществляют транспортировку питательных веществ к клеточным элементам и отдачу растворенных продуктов обмена обратно в кровь. Другими словами, соединительная ткань является важнейшим действенным физико-химическим фактором водного обмена, чрезвычайно чувствительным к изменениям содержания ионов в окружающих тканях (кровь или органы) и в свою очередь на эту среду влияющим (см. также Гидрофилия.,, гидрофильность). Многочисленные наблюдения над стойкостью эритроцитов в растворах нейтральных солей и над тканевыми культурами свидетельствуют, что набухание и сморщивание клеток находится в зависимости в первую очередь от осмотического давления окружающей pix среды. Однако в этом обмене с окружающей средой принимает участие не вся вода, входящая в состав клетки, а лишь та подвижная ее часть, к-рая играет роль растворителя, конституционная же вода, как указывалось, отдается протоплазмой с боль-щим трудом и то лишь перед гибелью клеточной структуры. Поэтому и степень набухания клеток не находится в прямом соотношении с концентрацией окружающего раствора (resp. тканевых соков). Кроме того на поверхности самой клетки имеют место сложные процессы адсорпции и изменения поверхностного натяжения, обусловливаю-I щего проницаемость клеточных оболочек пли их пограничных слоев. Эти оболочки сами по себе могут благоприятствовать или препятствовать осуществлению осмотических процессов в клетке и обмену воды в ней. Расшифрование подобных влияний в наст. время затруднено хотя бы потому, что мало известны все те физ.-хим. фазы и состояния, в к-рых находится протоплазма клеток и в частности ее пограничный слой, а также, какими силами удерживается вода клетками. Подчиняясь осмотическим влияниям, клеточная протоплазма, несущая сама определенный электрический заряд, но не способная к диффузии, участвует в т. н. аномальном осмосе, т. е. создает неодинаковую концентрацию в окружающей жидкости и внутри клетки для целого ряда анионов (СГ, НСОз). Другими словами, распределение этих ионов по обе стороны клеточной оболочки—между тканевой жидкостью и содержимым клетки—происходит неравномерно по законам т. н. равновесия Доннана (см. Доннана равновесие). Водный баланс организма складывается из воды, вводимой с пищей или образующейся в результате жизнедеятельности органов и тканей, с одной стороны, и воды, выводимой при помощи мочеотделения, перспирации (через легкие, кожу),—с другой. Введение воды в организм регулируется особым физиол. ощущением—жаждой, степень к-рой обусловлена в первую очередь содержанием осмотически активных веществ (resp. молекулярной концентрацией) в крови и тканевых жидкостях (см. Жажда). Что же касается выделения воды, то оно в физиоло-гич. условиях находится в строгом соответствии с количеством вводимой воды. При обильном питье регуляция эта осуществляется быстро (гл. обр. почками). Наступающий диурез является результатом тех сдвигов, к-рые происходят в водно-солевом хозяйстве организма. Сдвиги эти подчас неуловимы и не всегда относятся к изменениям состава крови; так, степень мочеотделения не находится в прямой зависимости от разжижения крови. Наряду с почками однако и экстраренальное выделение воды может оказать существенное влияние на водное хозяйство * организма, что имеет место гл. обр. при высокой внешней t° (климат, условия, работа в горячем цеху) или лихорадке. Испарение воды с сравнительно большой поверхности тела играет роль терморегулятора, способствующего при соответствующих условиях поддержанию t° тела на определенном уровне. Вместе с тем необходимо подчеркнуть, что при избытке воды в организме экстраренальное выделение и при нормальной t° окружающей среды резко нарастает, а в пат. условиях, например при гипертиреозе (в особенности экспериментальном), роль экстраренального выделения в водном балансе резко повышается, доходя до 50% всей выделяемой воды. Однако уже в норм, условиях количество воды, удаляемой экстраре-нальным путем, может доходить у человека до 800 — 900 см3 в сутки. Из этого количества 500—600 смъ выделяется при помощи кожной перспирации или потоотделения, а остальное количество в виде водяных паров выдыхаемого воздуха. Кстати сказать, прово- димое в отношении водного баланса качественное различие между перспирацией и потоотделением мало обосновано, т. к. первая осуществляется теми же потовыми железами, и только лишь невидима для невооруженного глаза. Отдача воды кожей дает более обширные возможности для регулирования водного хозяйства, нежели дыхание, т. к. кожная перспирация находится в прямой зависимости от чрезвычайно чувствительного к температурным колебаниям, ля-бильного и поддающегося тонкой регуляции иннервационного аппарата. Количество же воды в выдыхаемом воздухе, имеющем довольно постоянную температуру и следовательно стойкое напряжение водяных паров, подвержено значительно меньшим колебаниям. Способность к удержанию водного хозяйства на определенном уровне, resp. к выравниванию водного баланса, в физиол. условиях поддается изучению при помощи т. н. «водной пробы». При введении 1 сОО ом3 воды последняя с избытком выделяется в течение 3-—4 часов, причем ренальным путем выводится 80—90% у взрослых и 70—80% у детей (см. также обмен веществ у детей и Почки,—фнкц. диагностика). Результаты водной пробы находятся однако в теснейшей зависимости от предварительного питания и в частности от предварительного насыщения тканей водой и солями. Так, у кроликов, вскармливаемых «сухой» пищей (например овсом), водная проба выпадает значительно медленнее, нежели у питаемых богатой водой морковью (Siebeck). Все это в еще большей степени относится к человеку, употребляющему в пищу чрезвычайно разнообразное количество жидкости и NaCl (от 5 до 20 г в сутки). Из входящих в ее состав ионов Na+ и СГ первому принадлежит несомненно гидропигенное свойство, т. к. и другие соли, содержащие катионы Na+, также способствуют задержке воды (NaHC03,Na2HP04). Странным образом противоположное (мочегонное) действие оказывают несущие также положительный заряд катионы К и Са, их соли и содержащие их пищевые вещества (картофель, плоды). Следовательно характер предварительного питания не только по содержанию в нем воды, но и по минеральному его составу оказывает решающее влияние на течение водного обмена. Все это лишь свидетельствует, что водный обмен не должен быть рассматриваем как самостоятельно протекающий процесс, в частности вне связи с минеральным обменом. Иногда в эксперименте (деминерализация, искусственная полидипсия, перегревание), а также в пат. условиях, можно наблюдать развитие длительного отрицательного водного баланса; в таких случаях наступает обезвоживание (см.), сопровождающееся тяжелыми явлениями вплоть до симптомов резкой интоксикации. Не менее тяжелую картину представляют случаи с положительным водным балансом; обычно это— состояния, наступившие вслед за резкой и быстрой задержкой значительных количеств воды в организме. Подобные условия могут быть созданы введением больших ко- личеств жидкости с последующим впрыскиванием питуитрина (Вовси, Ицексон и Вагон). Наблюдаемая при этом картина «водного отравления» (Rowntree) характеризуется иногда чрезвычайно грозными симптомами (дрожь, беспокойство, бледность, тошнота и т. п.). В физиолог, же условиях в организме беспрерывно и чрезвычайно быстро происходит выравнивание водного баланса, создающее ту относительную устойчивость водного хозяйства, которая является предпосылкой нормального течения всех процессов промежуточного обмена. Подобная регулировка осуществляется в первую очередь, разумеется, центральным путем, что было подтверждено экспериментально при помощи т. н. «водных» и «солевых» уколов. Аналогично Клод Бернаровскому «сахарному» уколу, сопровождающемуся наряду с гликозурией также и полиурией, Мейер и Юнгман, а также Лешке (Meyer, Jungmann, Leschke) показали, что укол в определенные участки серого бугра и промежуточного мозга влечет за собой усиленное мочеотделение. Оно может сопровождаться одновременным повышенным выделением поваренной соли («солевой» укол) или же ограничиваться выделением жидкой, бедной солью мочи («водный» укол). Центрипеталь-ные импульсы этими центрами получаются повидимому либо непосредственно из тканей (см. Жаоюда) либо же через гормональный аппарат, влияние которого на функцию водорегулирующих центров исключительно велико (см. ниже). Из желез внутренней секреции задней доле гипофиза, щитовидной железе и поджелудочной железе принадлежит решающая роль в регуляции водного хозяйства организма. Экстракты задней доли гипофиза (питуитрин и его аналоги) обладают, как известно, резким антидиуретическим действием, причем остается неясным вопрос, обусловлен ли этот эффект специальным гормоном или же дело идет об одном из свойств единого гипофизарного инкрета (см. Гипофиз). Антидиуретическое действие питуитрина особенно рельефно сказывается при обильном диурезе: после водной нагрузки, после экстирпации гипофиза у собак или при diabetes insipidus (см. Диабет несахарный). В крови после инъекции питуитрина как правило отмечается более длительное разжижение ее. а в тканях накопление жидкости (resp. повышение веса тела). Мейер и Мейер-Биш (Meyer, Meyer-Bisch) показали также, что наблюдаемого обычно усиленного поступления лимфы в грудной проток из печени при введении т. н. lym-phagoga первого порядка (см. Лимфа) не происходит, если одновременно с этими веществами впрыснуть питуитрин. В последнем случае из фистулы грудного протока поступает малое количество густой лимфы. Наряду с этим имеются многочисленные наблюдения, что и в самих почках действие питуитрина сказывается преходящим, но все же резким сужением почечных сосудов и уменьшением почечного кровообращения. Поэтому многие исследователи склонны связывать падение диуреза с влиянием питуитрина непосредственно на почки. Во всяком случае антидиуретическое влияние питуитрина не является самодовлеющим и осуществляется через центральную нервную систему, а не непосредственным воздействием на исполнительные органы, т. к. при перерезке пп. splanchnici, при разрушении серых бугро'в или наконец при выключении мозговой коры действие это либо выпадает либо резко уменьшается. Механизм действия и точка приложения гормона щитовидной железы, также участвующей в водном или вернее в водно-солевом обмене, совершенно отличны. Вещество щитовидной железы (тиреоидин и его аналоги), а также и ее гормон тироксин обладают ясно выраженным обезвоживающим действием. При длительном приеме внутрь тиреоидина наблюдается обычно падение веса тела при почти нормальном диурезе. Это похудание обусловливается больше повышением экстраренального выделения воды, нежели, усилением мочеотделения. Особенно эффективно обезвоживающее действие тироксина при наличии предварительной задержки в тканях воды и поваренной соли, в частности при микседеме, при отеках у декомпенсированных сердечных б-ных, при нефрозе, при ожирении. Благодаря этому имеются все основания причислить гормон щитовидной железы к разряду водо- и солегонных веществ с точкой приложения в тканях. В частности похудание, наблюдаемое при приемах тиреоидина у тучных, сопровождается не столько повышением у них основного обмена, сколько удалением из организма значительного количества солей и воды. Инкреты инсуляр-ного аппарата поджелудочной железы также способствуют задержке воды в организме. Ретенирующее влияние инулина, в отличие от функции гипофиза, сказывается однако лишь в тех случаях, когда вместе с водой в организм вводится известное количество Na-солей и углеводов. Повидимому под влиянием инсулина усиление процесса глюконеогенеза сопровождается задержкой воды в печени. Точно также и уси-' ленное введение натриевых солей, особенно щелочно реагирующих (бикарбонаты, двуосновные фосфаты), уже само по себе способствует задержке воды в тканях и даже благоприятствует образованию отеков. Одновременное же введение инсулина лишь ускоряет и усиливает этот процесс. Патология водног > обмена. В пат. усло-•виях нарушения водного хозяйства имеют место при самых различных заболеваниях. Схематически все эти отклонения можно свести к страданиям или симптомам с положительным или отрицательным водным балансом. Отрицательный баланс наблюдается гл. обр. в детском возрасте при т. н. эксикозах (см.), атрофиях, при резких поносах (в частности при холере и детской диспепсии), при кахексиях, в начальном периоде голодания, при перегревании и потогонных процедурах. Во всех этих случаях причиной отрицательного водного баланса является преимущественно экстра-ренальное выделение воды. Избыточная потеря организмом жидкости имеет место так-I же при полиуриях различного происхожде- ния, из к-рых на первое место должно быть поставлено мочеизнурение сахарное и несахарное. При всех этих заболеваниях падают ■эластичность и жизненный тур гор в первую очередь тех тканей, к-рые являются водным депо организма, как-то: соединительной и мышечной ткани; наряду с ними обезвоживается также и кровь. Величина сухого остатка крови (количество белков и солей) достигает высоких цифр. Содержание белков плазмы доходит до 10%, a NaCl—до 650 мг%. Соответственно понижается также точка замерзания крови. Резкие потери в весе, наблюдающиеся при этих страданиях, обусловлены в первую очередь потерей значительного количества тканевой межклеточной воды. Однако тканевое обезвоживание не проходит бесследно для коллоидов клеточной протоплазмы, в особенности паренхиматозных органов, следствием чего являются наблюдаемый обычно во втором периоде обезвоживания усиленный белковый распад ■с выделением избыточных количеств азота с мочой (отрицательный азотистый баланс). Продолжающееся мочеотделение при высоком процентном содержании поваренной •соли и азота в моче создает предпосылки для возникновения основного симптома, наблюдаемого при отрицательном водном •балансе,—жажды. В случаях с более продолжительной или резко выраженной потерей воды к жажде присоединяются двигательное беспокойство, чувство тяжести в голове, ощущение жара, боли в конечностях, •а иногда и псих, возбуждение. Описываемые признаки интоксикации едва ли относятся к одной лишь потере воды, а скорее близки к весьма характерной картине, описанной ■Фишлером (Fischler) под названием мясной интоксикации и наблюдаемой при поступлении в кровь продуктов расщепления белка. Интересно отметить, что все упомянутые ■симптомы могут возникнуть также при скоплении значительных количеств жидкости внутри организма, т. е. при видимо положительном водном балансе, напр. при нарастании асцита у больных цирозом печени. В подобных случаях эти явления обусловлены патогенетически тем же обезвоживанием ткани, resp. сгущением крови, имеющим здесь место так же, как и при значительных потерях воды. Полиурии, приводящие как правило к отрицательному водному балансу, могут происходить от самых разнообразных причин. Наиболее часты и общеизвестны полиурии при диабете ■сахарном и несахарном, в начале почечной недостаточности, при т. н. первичной полидипсии и др. (см. Полиурия, Нефрит, Не-фросклероз, Полидипсия). Показателем возникновения положительного водного баланса и признаком задержки воды в организме естественно было бы считать олигурию. Однако опыт учит, что при т. н. первичных олигу-риях, обусловленных чаще всего нервными или эндокринными воздействиями (см. Оли-гурия), водный баланс выравнивается экстра-ренальным выделением воды и задержки ее т. о. не наступает. Поэтому к состояниям, характеризующим собой истинный положительный баланс, относятся лишь те, к-рые сопровождаются быстрым нарастанием общего веса б-ного с олигурией или без нее. Наиболее выражена задержка воды в органах в период развития или нарастания видимых или скрытых отеков. Последние возникают преимущественно вследствие нарушения одного или нескольких из перечисленных факторов, регулирующих промежуточный обмен воды в организме. Так. застойные отеки связаны чаще всего с повышением гидростатического давдения в венозной системе и падением наряду с этим проницаемости капилярного эндотелия для воды, обусловленным его кислородным голоданием (Landis). Почечные отеки, в особенности нефротические, имеют очевидно в основе своей иной генез и связаны в первую очередь с понижением количества белков в плазме, изменением дисперсности их частиц, сдвигом альбумино-глобулинового кое-фициента и следовательно падением коллой-до-осмотического давления белков, т. е. с уменьшением той силы, с к-рой белки удерживают свою воду. Наконец сдвиги в активной реакции тканей, накопление в них нек-рых кат- и анионов (Na+,HC07), также влекут за собой значительную задержку воды в тканях (воспалительные, голодные, алиментарные, солевые отеки). (Подробнее см. Отек.) Преимущественно тканевой характер носит очевидно также значительная задержка воды, наблюдаемая в начальном периоде различных инфекц. заболеваний (например при крупозной пневмонии). Лейден (Leyden) на большом материале показал, что потеря воды в лихорадочном периоде-в 2 раза меньше, нежели в посткритическом. Предположение, что решающая роль в этой задержке принадлежит лихорадке, едва ли обосновано, так как подобная картина наблюдается и после туберкулинизации. Несомненно лишь, что задержка воды в этих случаях происходит параллельно, а быть может и обусловлена резкой задержкой поваренной соли и усилением белкового распада, наблюдаемыми при этих состояниях. О патологической анатомии изменений при нарушении водного обмена— см. Вакуольное перерождение. Отечная болезнь, Отек. м. вовси. ІX. Обмен веществ у детей. О. в. и обмен энергии у детей, особенно раннего возраста, по сравнению с обменом взрослых протекает во многих отношениях совершенно своеобразно. Для получения достаточно точных результатов при изучении О. в. у детей необходимо кроме пунктуального соблюдения обычных общих правил исследования обмена у взрослых учитывать еще следующие основные моменты: 1) исследуемая группа детей должна быть по возможности однородна в отношении состояния здоровья, конституции, возраста и способа вскармливания; 2) условия наблюдения во всех сравниваемых между собой случаях должны быть совершенно одинаковы; 3) продолжительность наблюдения должна быть не менее 5—7 дней; подготовительный период с нагрузкой ребенка основной диетой должен быть достаточно продолжителен (3—4 дня); 4) необходимо учитывать 752" влияние времени года, освещения, богатства солнечного спектра ультрафиолетовыми лучами и других метеорологич. моментов; 5) особое внимание должно быть обращено на тщательное собирание мочи и стула, что у детей раннего возраста представляет особые трудности и требует специальной методики. Для этой цели предложено много различных способ^, из которых лучшими надо призййть способ Бендикс - Финкель-штейна (ВепШх, Finkelstein) и способ Бен-жамина (Benjamin). Положение ребенка и мочеприемника при собирании суточного количества мочи—см. вт. XIX (ст. 107, рис. 4). Стул собирается на резиновую подстилку. Для изучения газообмена у детей имеются специальные камеры (при аппаратуре Бенедикта и Книппинга) (Benedict, Knipping). Для исследования крови приходится пользоваться гл. обр. микрометодами. В остальном изучение различных видов О. в. у детей обычно проводится с помощью тех же методов, что и у взрослых. Обмен энергии (основной обмен). Энергия, получаемая ребенком и оцениваемая калоражем принимаемой им пищи, расходуется на поддержание жизни натощак в состоянии покоя (основной обмен), на рост и нарастание массы тела и на мышечную деятельность. Гельмрейх (Helmreich) дает след. примерное распределение энергии у ребенка и взрослого при умеренной пище: табл. 26. Распределение энергии На основной обмен ...... На рост и отложение вещества............... На специфически динамическое действие ............ На- работу для передвижения тела............... На потери экскретами..... У ребенка (в %) У взрослого (в %) 15 0—5 15 5—10 Основной обмен у ребенка зависит не только от его живой массы, но также и от индивидуально различной у отдельных субъектов интенсивности обмена веществ в клеткахТ*У одного и того же субъекта основной обмен отличается удивительным постоянством и колеблется в пределах около 10%. Между величиной основного обмена у ребенка и его возрастом, весом и поверхностью тела существует известная зависимость. Для определения поверхности тела у грудного ребенка пригодна формула Лиссауера (Lissauer): со (поверхность тела) = 10,3[^вес2; для детей более старших константа 10,3 должна заменяться другими, различными в зависимости от веса ребенка; величина константы для мальчиков колеблется от 10 до 11,5, для девочек от 10,1 до 11,1 (Talbot). Формула Лиссауера пригодна гл. обр. для нормальных детей. Более точной является формула Дюбуа (du Bois): поверхность тела=вес °'425 х х рост°'725х 71,84. Бенедикт и Тальбот для вычисления основного обмена у новорожденных (весом до 6 кг) предлагают след. формулу: суточная выработка калорий = = 12,65 х длину (в см) х 10,3 i^bgc2 (в кг). У детей периода новоро*кд"енности основной обмен очень низок и приблизительно пропорционален их весу. По исследованиям ряда авторов (Бенедикт, Тальбот, Bailey, Murlin) суточное расходование энергии у новорожденного с весом в 31/2 кг составляет приблизительно 146 кал., или 42 кал. на 1 кг веса; совершенно такие же соотношения между расходом энергии и весом получаются и у детей первого дня жизни с более низким и более высоким первоначальным весом; ко 2—3 дню жизни расход энергии на 1 кг веса повышается до 48 кал. и, возрастая постепенно в дальнейшем, достигает к lyj2 годам максимальных цифр (50—60 кал. на 1 кг), а затем начинает снижаться. У мальчиков в возрасте от 2 до 3 лет он падает до 52 кал., к 6—7 годам до 42 кал., к 10—11 годам до 38 кал. и к 12—13 годам до 34 кал. на 1 кг веса (Бенедикт). Т. о. можно считать бесспорным, что основной обмен, рассчитанный на единицу веса, гораздо выше в раннем возрасте, чем у детей более старших. Различие между основным обменом у мальчиков и девочек начинает выявляться приблизительно с 1— V-j2 лет; с этого возраста и до периода полового созревания расход энергии у девочек несколько меньше, чем у мальчиков, затем кривая девочек обгоняет кривую мальчиков и лишь с окончанием полового созревания у мужчин снова восстанавливается преобладание обмена. Это половое различие, надо думать, зависит от меньшего у девочек количества энергетически активной ткани, но зато большего количества жировой ткани. Дети упитанные, ткани к-рых богаты жиром и водой, дают более низкие цифры основного обмена, чем дети худощавые. В период полового созревания в нек-рых случаях повидимому наступает повышение основного обмена. Возрастные изменения основного обмена у детей, рассчитанные не на возраст и единицу веса, а на единицу поверхности тела,. характеризуются следующими цифрами: у новорожденного около 612 кал. на 1 м'% к концу года—около 1 100, к 10 годам— 1 000 и к 13 годам—950. Дыхательный кое-фициент (см.) в период новорожденности претерпевает следующие изменения: в первый день 0,80, к третьему дню падает до 0,73 и затем к 5—6 дню достигает 0,81 — 0,82.—«Специфически динамическое' действие» пищи у детей раннего возраста несколько иное, чем у детей более старших и взрослых. По исследованиям Гельмрейха введение с пищей молока у маленьких детей почти не вызывает повышения основного обмена, тогда как у больших детей нарастание расхода калорий довольно значительно и достигает приблизительно 10%. В рериод полового созревания по данным Гетхе (Gottche) специфически динамическое действие в 75% случаев было сильно понижено и в большинстве случаев (50%) сопровождалось повышением основного обмена. Закономерность этой реакции полового созревания (Pubertatsreaction), как называет ее Гетхе, не может считаться окончательно доказанной. Не подлежит сомнению, что состояние обмена энергии в растущем орга- низме находится под большим влиянием со стороны эндокринного аппарата. Азотистый обмен. Белки пищи всасываются у детей преимущественно в виде аминокислот и отчасти в виде полипептидов. Прохождение нативных белков через нормальную кишечную стенку повиди-мому возможно только у детей периода но-ворожденности, когда некоторые белковые тела могут непосредственно проникать через неповрежденную слизистую оболочку. О дальнейшей судьбе в организме ребенка всосавшихся в кишечнике продуктов белкового расщепления известно очень мало. Самый процесс всасывания белков в кишечнике, особенно у детей раннего возраста, происходит очень совершенно и почти не находится в зависимости от возраста ребенка и способа его вскармливания; у детей на грудном вскармливании всасывается от 70 % до 90% (в среднем около 80%) введенных белков, у детей на искусственном питании лишь несколько меньше. Повышенное введение с пищей белков, как показывают исследования Финкелынтейна и Ионас (Fin-kelstein, Jonas), не ухудшает процесса их всасывания. Что касается усвоения всосавшихся аминокислот и полипептидов, то по более старым наблюдениям (Langstein, Meyer, Rietschel и др.) процент используемого азота пищи колеблется в зависимости от возраста ребенка и споссба его вскармливания, тогда как количество белка, задерживающегося в организме, находится в зависимости от возраста ребенка и почти не зависит от количества белков пищи. Однако новейшие исследования Ромингера и Мейе-ра (Rominger, Meyer H.) показывают, что и задержка азота в организме здорового грудного ребенка в значительной мере зависит от количества вводимого белка; детский организм в противоположность организму взрослого обладает выраженной способностью к повышенному накоплению белка, благодаря чему ткани ребенка на искусственном вскармливании несколько богаче белками, чем ткани ребенка, получающего только грудь. Несколько своеобразные отношения отмечаются у детей периода новорожденное™, у к-рых по.наблюдениям Ланг-штейна, Нимана и Бирка (Niemann, Birk) отмечается отрицательное азотистое равновесие, быстро и достаточно совершенно выравнивающееся при естественном вскармливании молозивом и значительно затягивающееся при искусственном питании. Соотношение между количеством белка в пище и количеством азота, выделяемого с испражнениями, остается б. или м. постоянным; при обычной белковой нагрузке у ребенка со стулом выделяется около 10% введенных белков. По данным Пинтоцци (Pintozzi) количество азота в .испражнениях может служить мерилом всосавшегося азота, т. к. только сравнительно небольшая часть азота стула относится за счет азота пищеварительных соков, изливающихся в жед.-киш. тракт, и совершенно ничтожное количество за счет выделяемых кишечником конечных продуктов белкового обмена. Главная масса последних, так же как и у взрослого, выделяется с мочой, но распределяют- ся они в ней совершенно своеобразно. Общее количество азота мочи относительно очень велико у детей периода новорожден-ности, потом оно несколько уменьшается с тем, чтобы в дальнейшем снова начать нарастать приблизительно параллельно количеству белков, вводимых с пищей. У грудных детей независимо от способа вскармливания около 40—60% азота, введенного с пищей, снова выделяется с мочой. Суточное количество азота мочи у новорожденного равно в среднем около 0,5 г, к концу первого года жизни—1,5 г, в возрасте 6—7 лет— 7,5 гик концу периода детства около 10 г, что дает при пересчете на 1 %г веса тела: 0,12—0,16 у грудного ребенка, 0,45 у ребенка 6—7 лет и 0,31 у детей периода полового созревания. Нек-рое общее представление о распределении азота в моче грудного ребенка может дать следующий пример Каммерера (Kammerer): из 5,52 г общего количества азота в моче ребенка, находившегося под наблюдением, 4,8 г приходилось на мочевину, 0,29 г—на аммиак, 0,065 г—на мочевую к-ту и 0,013 г—на пуриновые основания. По данным последнего автора у ребенка, получающего грудь, за счет мочевины приходится около 75—80% общего азота мочи, у рожковых детей—около 84% и у детей более старшего возраста—от 90% до 94%; возрастная кривая выделения мочевины проделывает приблизительно такие же колебания, как и кривая общего количества азота. Мочевой к-ты в период новорожденности выделяется приблизительно в 5 раз больше, чем у взрослого, и в два раза больше, чем у детей более старших, у к-рых на 1 кг веса ее выделяется от 0,018 до 0,027 г pro die; эта величина мало меняется в зависимости от возраста ребенка, но в значительной мере зависит от принимаемой им пищи (Gopport). Пуриновые основания выделяются параллельно количеству мочевой к-ты (Niemann), но в моче новорожденных они содержатся только в виде следов. За счет аммиака мочи у детей выделяется больше N, чем у взрослых; так, у новорожденных эта величина составляет 9,5—19,5% общего количества мочевого азота (Reuss, Келлер), в грудном возрасте—8%, к 3—4 годам—7% и в 13— 14 лет—6%. Процентное отношение азота аммиака к общему количеству азота, т. е. так наз. аммиачный коеф. мочи, колеблется у детей в широких пределах (в норме от 3 до 10), изменяясь в значительной мере в зависимости от состояния кислотно-щелочного равновесия в организме, нарастая при состоянии ацидоза и уменьшаясь при сдвигах в сторону алкалеза, и является т. о. довольно чутким показателем правильности течения общего обмена. Азот аминокислот дает высокие цифры в первые дни жизни; особенно много его в моче недоносков (до 9,5—25% по Gebel'ro); в грудном периоде величина эта колеблется от 2,7% до 4%, т. е. в 2—2V2 раза больше, чем у детей старших, у к-рых за счет аминокислот выделяется всего около 1,6—1,8% общего азота мочи. На выделении с мочой конечных продуктов азотистого обмена кроме возраста ребенка 7бв. сказываются и его конституциональные особенности (Маторина, Мац, Тур). Распределение азота в моче зависит от состояния окислительной способности организма ребенка; в результате недостаточности ее у новорожденного и является повышенное выделение с мочой большого количества продуктов неполного азотистого обмена, что свидетельствует о неприспособленности организма к полному расщеплению белков и следовательно о невозможности достаточно совершенного использования белков пиши. Энергия окислительных процессов с возрастом усиливается, достигает оптимальных условий к 5—6 годам и затем снова снижается к периоду полового созревания. Эти особенности азотистого обмена, а также основное свойства детского организма— энергичное течение процессов роста, возможное только при наличии большого количества пластического материала,—делают необходимым введение детям с пищей относительно больших количеств белков. У детей, искусственно вскармливаемых, наряду с большой задержкой в организме вводимого азота отмечается также и усиленное выделение его с мочой (Ромингер и Мейер), и следовательно пищевой рацион таких детей должен быть богаче белками, чем для детей, вскармливаемых только грудью. В связи с своеобразием интермедиарного обмена белков у детей находится повышенное выделение у них с мочой органических к-т: у недоносков и новорожденных в сутки—10,1 см3 '7ю кислоты, у детей грудного и дошкольного возраста —12,5, тогда как у взрослых только 8,2 см2 на 1 кг веса (Hot-tinger). По мнению большинства авторов за счет белков у детей должно покрываться приблизительно от 10% до 15% общего количества калорий. Потребность в белках у ребенка первого года жизни в среднем около 2,5 г на 1 кг веса (у грудного от 1,5 г до 2 г, у рожкового от 3 г до 4 г), в возрасте от 1х/г до 5 лет несколько повышена и в среднем равна 3—3,5 г, к 8—10 годам падает до 2,5 г и к 12—15 годам до 1,5—2 г; при наличии известных показаний количество вводимых белков может быть как увеличиваемо, так и уменьшаемо в довольно широких пределах без нарушения при этом нормального течения процессов обмена и правильности общего развития. По данным многих немецких авторов потребность ребенка в белках значительно меньше. Для правильного развития детского организма не столь важно абсолютное количество вводимых белков, как их полноценность, определяемая содержанием и характером аминокислот. К жизненно необходимым для ребенка аминокислотам надо отнести триптофан, лизин, тирозин и цистин, к менее важным, но в некоторых случаях безусловно необходимым,—аргинин, гистидин, аспарагин и глю-таминовую к-ту. Недостаточное введение указанных аминокислот неизбежно ведет к развитию дистрофии, а полное отсутствие несовместимо с жизнью. Потребность в триптофане в грудном периоде—0,058—0,1, в более старшем—0,055—0,063 на 1 кг веса. Практически достаточно, чтобы около 50% вводимых белков было животного происхождения. Для правильного усвоения белков необходимо известное соотношение (корреляция) с другими основными ингредиентами пищи—жирами, углеводами и водой. Избыток вводимых белков не улучшает, а иногда повидимому даже ухудшает использование азота, и ведет к состоянию относительного ацидоза; избыточное введение жиров по одним авторам (Freund, Neumann) ухудшает всасывание и задержку азота, по другим (Пинтоцци) не оказывает никакого влияния; углеводы, по Келлеру (Keller), улучшают азотистый баланс, а по Пинтоцци, несколько нарушают всасывание белков и уменьшают потребность в них. Умеренное преобладание в пище щелочных эквивалентов над кислотными улучшает усвоение белков. Недостаточное введение воды благодаря выраженному специфически динамическому действию белков может вести к гипертермии (Eiweissfieber). Специфически динамическое действие белков с возрастом ребенка, т. е. с увеличением поверхности его тела, нарастает; у маленьких детей оно меньше, чем у более старших: у последних меньше, чем у взрослых (Helmreich, A. Seifert); у мальчиков больше, чем у девочек. Максимум специфически динамического действия белков отмечается через 3 ч. после приема пищи и колеблется между 9,2% и 31,4% цифры натощак. Жировой обмен у детей также отличается известным своеобразием. Можно считать установленным, что даже у детей раннего возраста расщепление жиров начинается уже в желудке под влиянием липазы желудочного сока; существенное значение для переваривания жиров у грудного ребенка имеет наличие большого количества липазы в женском молоке. Процесс расщепления и механизм всасывания жиров у детей в основном протекает так же, как и у взрослых. По Орглеру, Келлеру, Э. и Ф. Мюллерам и по новейшим данным Поттера (Potter), у ребенка на грудном вскармливании всасывается около 98%, у ребенка рожкового—около 95%, у детей более старших—от 95% до97% жиров, вводимых с пищей. Большая часть расщепленных и всосавшихся в кишечнике жиров, поступая через воротную вену и печень в общее кровяное русло, ведет к появлению т. н. пищеварительной гиперлипемии; последняя нарастает в течение всего процесса пищеварения, достигая максимума при коровьем молоке через 3 часа, при женском молоке— несколько позже, и держится в первом случае около 5 часов и во втором около 9 часов. В сыворотке крови детей натощак содержится 170—200 мг% фосфатидов, 140—170 мг% холестерина и около 400 мг% жирных кислот. В крови новорожденного липоидов значительно меньше, но унте к 1—11/2 месяцам количество их увеличивается на 50—75% и затем медленно, но постепенно нарастает в течение первых лет жизни. В возрасте от 3 до 11 лет общее количество липоидов в среднем равно 700 мг%, что почти соответствует содержанию их у взрослого. Способ вскармливания ребенка заметным образом не отражается на количестве липоидов кро- ви. Неиспользованные организмом жиры в количестве около 5—10% (по наблюдениям Фрейнда до 14—18%) выделяются со стулом в виде нейтрального жира, нерастворимых в воде свободных жирных к-т, щелочных (растворимых в воде) и щелочноземельных мыл. Распределение их в кале грудных и рожковых детей несколько различно. У нормального грудного ребенка жиры испражнений содержат нейтрального жира 10—30%, свободных жирных кислот—10%, щелочных мыл—40%, щелочноземельных— 20%, холестерина—0,8% и незначительное количество лецитина; у искусственно вскармливаемого ребенка нейтрального жира около 35%, свободных жирных к-т—-до 40%, щелочных мыл—10% и щелочноземельных—■ 13%. У грудного ребенка со стулом выделяется холестерина больше, чем вводится с пищей, что указывает на возможность его синтеза в растущем организме; однако Кнау-ер ;Knauer) это предположение считает маловероятным. Распределение жира в стуле меняется в зависимости от состава пищи и состояния кишечной перистальтики. Всосавшиеся жиры отчасти сгорают в организме и т. о. являются источником образования энергии, отчасти, отлагаясь в тканях, гл. образом в подкожной клетчатке и брыжейке, образуют резервное жировое депо. Ж'лр грудного ребенка беден жидкой олеиновой к-той; с возрастом количество ее постепенно возрастает. Растущий организм сравнительно долгое время может развиваться без жиров на одних белках и углеводах, а последние могут быть исходным материалом для образования жира, отлагающегося в тканях. Однако эта возможность замены жиров изодинамическими количествами углеводов свидетельствует только о большой приспособляемости детского организма, но не дает права на заключение, делаемое нек-рыми авторами (Pirquet, Greer), что жиры не являются безусловно необходимыми для организма. Во-первых жир, образующийся в организме из углеводов, отличается по своему составу от жира при нормальном питании и во-вторых при обезжиренной, но в остальном калорийно достаточной пище рано или поздно у ребенка развиваются явления авитшиноз! (Bloch, Wagner), появляется ксерофтальмия и резко падают иммунные силы организма (Weigert). Нормальная потребность в жирах в грудном возрасте относительно велика, обычно колеблется от 3 до 5 г, в среднем же равна 4 г на 1 кг веса, после года—3,5 г, от 6 до 12 лет—3 г и в период полового созревания— 2—2,5 г. Если, с одной стороны, детский организм повидишму может правильно развиваться и при значительно меньшем количестве вводимых жиров, то, с другой стороны, количество их при наличии известных показаний может быть значител но повышено (напр. до 6 г и более на 1 кг веса в грудном возрасте). Однако избыточное введение жиров не безразлично для организма и легко ведет к развитию состояния истинного ацидоза. Потребность в жирах определяется не только количеством их, но и качеством; полноценность жиров зависит от содержания в них липоидов и витаминов, что особенно должно учитываться при питании детей раннего возраста. Правильное течение жирового обмена возможно лишь при надлежащей корреляции жиров с другими питательными началами; у грудного ребенка за счет жиров должно покрываться до 50% всех вводимых калорий, у старших детей всего лишь 25—30%. Углеводный обмен. Обмен углеводов у ребенка, подчиняясь в общем основным законам, регулирующим его у взрослого, отличается рядом характерных особенностей. Выключение углеводов из пищи вызывает у ребенка понижение темлерату-ры тела; этот факт подтверждает громадное значение углеводов для теплообразования и терморегуляции в организме ребенка. Вопрос о возможности повышения температуры тела у детей под влиянием избыточного введения сахара пока не может считаться окончательно решенным. Углеводы всасываются глав шм обр гзом в виде моносахаридов (глюкоза, левулеза, галактоза), хотя не исключена возможность всасывания и декстринов. В крови здоровых грудных детей количество сахара натощак колеблется от 0,07% до 0,09%, у детей более старших от 0,08% до 0,1% и наконец у детей 12— 14 лет от 0,09% до 0,12%. В период пищеварения количество сахара в крови повышается до 0,16—0,2%; эта гипергликемия быстро выравнивается, так как часть сахара окисляется, используясь для чисто динамических целей, часть откладывается гл. обр. в печени и отчасти в мышцах в виде гликогена и наконец часть используется для образования жира. Способность детского организма ассимилировать вводимые с пищей углеводы имеет известный предел; если этот предел ассимиляции превзойден и количество сахара в крови б. или м. стойко нарастает до 0,18—0,2%, то организм выравнивает нарушенное углеводное равновесие путем выведения избытка сахара с мочой. Выносливость ребенка к углеводам подвержена значительным индивидуальным колебаниям; она находится в зависимости от возраста ребенка и конституциональных особенностей его организма, она неодинакова к различным углеводам и вообще значительно больше, чем у взрослого. У грудного ребенка пищевая лактозурия наступает при введении 3,5 — 4 г молочного сахара на 1 кг веса; у взрослого для этого достаточно всего лишь 1 г. Тростниковый сахар имеет такой же предел ассимиляции, как и лактоза; для виноградного сахара и мальтозы граница выносливости лежит несколько выше, а левулеза имеет особенно низкий предел ассимиляции. По Ашенгейму (Aschen-heim), пищевая гликозурия у грудных детей наступает при введении на 1 кг веса 4 г галактозы, 8—12 г декстрозы, 4—5 г ле-вулрзы, 6—8 г лактозы и 6—8 г сахарозы. Эти абсолютные цифры имеют весьма незначительную ценность, они сильно меняются не только в зависимости от индивидуальных особенностей ребенка и состояния его жел.-киш. тракта, но и от той растворяющей среды, в к-рой эти углеводы вводятся. Всасывание сахара при женском молоке происходит быстрее, и гликемическая кривая поднимается раньше и падает круче, чем при цельном коровьем молоке. Водный раствор сахара вызывает крутой (максимум через 1/2—1 час) подъем и раннее угасание (через 2—3 часа) гликемической кривой. На гликемической кривой, как показывают исследования Стыриковича, отражаются во-первых состояние деятельности жел.-киш. тракта и печени (первый подъем кривой) и во-вторых особенности интермедиарного обмена (вторая половина кривой). Белки, жиры и углеводы, вводимые одновременно с сахаром, характерным образом изменяют гликемическую кривую,.что зависит от возникающих при этом изменений со стороны межуточного обмена. Своеобразие гликемич. реакций в раннем возрасте свидетельствует о напряженности и мобильности у детей углеводного обмена и о тесной связи его с межуточным обменом белков и жиров. В качестве одного из промежуточных продуктов интермедиарного обмена глюкозы в организме образуется молочная к-та. Гликоли-тическая способность организма, как показывают исследования Варбурга (Warburg), имеет большое значение для процесса роста. По Гьёрги, Бреме и Бради (Gyorgy, Brehme, Brahdy), в крови детей первой четверти года содержится в среднем 18,7 мг% молочной к-ты, у грудных детей более старших—13,8 мг%, тогда как у взрослых только 10,2 мг%. Часть углеводов, избегнувших всасывания, подвергается брожению под влиянием бактерий. Возникающие при брожении в кишечнике низшие жирные к-ты в малом количестве полезны и необходимы для организма: они возбуждают перистальтику кишечника, улучшают азотистый и минеральный обмен и являются т. о., как говорит Клоц (Klotz), катализаторами О. в. Наоборот, большие количества этих кислот действуют разрушительно, содействуя бактерийной инвазии в верхние отделы кишечника, подавляют обмен и ухудшают усвоение. Какая часть углеводов подвергается ферментативному расщеплению и всасывается и какая часть подвергается брожению—установить точно нельзя. В испражнениях здорового грудного ребенка углеводы (моносахариды) отсутствуют; при ускорении перистальтики появляются в испражнениях сахар и крахмал. Углеводы, как и белки, являются безусловно необходимыми для растущего организма и полностью не могут быть заменены изодинамическим количеством жиров. Даже кратковременное полное исключение их из пищи с трудом переносится организмом, в особенности если при этом вводится большое количество белков и жиров (наклонность к ацидозу). Необходимый для ребенка минимум углеводов повидимому индивидуально различен и в значительной мере зависит от количества и качества других пищевых ингредиентов, получаемых ребенком. Если этот минимум углеводов отсутствует в пище ребенка, то организм стремится пополнить его за счет расщепления белков. Недостаточное введение углеводов ведет к нарушению и водного обмена. Суточная потребность грудного ребенка в углеводах—около 10—12 з на 1 кг веса. В возрасте от 1 года до 5 лет в среднем около 10 г на 1 кг веса, а по нек-рым авторам даже и меньше; после 10 лет суточная потребность падает до 8—-9 г. Потребность в углеводах в абсолютных цифрах—около 120—130 г к концу первого года жизни, 150—160 з в 3 г., около 200 г в 6 лет, около 225—250 г к 10 годам и до 350—450 з к 14—15 годам. В первое время все потребности ребенка покрываются одним молочным сахаром, с 5 мес. появляется потребность в полисахаридах. Соотношение с другими основными ингредиентами пищи в грудном возрасте наиболее благоприятно, если оно соответствует таковому в женском молоке, т. е. белки относятся к жирам и к углеводам как 1 :2,8: 5,5, а жиры к углеводам как 1:2. При количестве белков, принятом за 1, отношения складываются так: 1:3,5:7. В более позднем возрасте эти отношения могут колебаться в более широких пределах, но желательно, чтобы за счет углеводов покрывалось до 35—60% всех калорий. Минеральный обмен в растущем организме является еще более сложным и во многих отношениях еще менее изученным, чем обмен органических веществ. Соли в экономике интенсивно развивающегося молодого организма играют весьма важную и притом крайне многообразную роль; длительное отсутствие даже одного основного иона несовместимо с жизнью и правильным развитием ребенка. Соли до известной степени предопределяют правильное течение водного, азотистого, жирового и углеводного обмена; без них не может совершаться построение новой ткани и возобновление старой, отмирающей в процессе работы. За счет солей поддерживается определенная концентрация водородных ионов, с нарушением к-рой меняется щелочно-кислотное | равновесие; самые незначительные наруше-| ния последнего неблагоприятно отражаются на нормальном течении всех жизненных процессов и нередко опасны для жизни ребенка. Сохранение осмотического давления, нормальное состояние коллоидов, поддержание известного равновесия в состоянии возбудимости нервной системы и нормальная ферментативная деятельность организма ребенка находятся в непосредственной зависимости от изменений со стороны минерального обмена. Для растущего организма особенно важны соли, принимающие участие в построении тканей. Если из пищи ребенка, вполне полноценной по своей калорийности, изъять минеральные вещества, наступает сперва задержка нарастания, а потом и падение весовой кривой. Всасывание вводимых с пищей солей происходит гл. образом в тонких кишках. Всасывание солей в кишечнике и задержка их в организме несколько различны в зависимости от возраста ребенка и способа его вскармливания. Эта способность уже достаточно хорошо выражена у новорожденного, удерживающего в организме за cvtkh, по Михельсу (Michels), 170—256 мг РА, 181—291 мг СаО и 95 мг С1. Длительные наблюдения Ромингера с сотрудниками дают следующие средние цифры, характеризующие минеральный обмен I у детей первого года жизни: Зид вскармливания При грудном вскармливании . При искусственном вскармливании ....... При смешанном вскармливании . Введено | t. a день Табл. 27 Задержалось в абсолют-! ТТТ)ПГТРП ном коли-! Б \?a3fe 1,07—1,5 0,37—0,72 2,46-5,8 ; 0,83—3,8 2,33—3,9 ; 1,12—1,3 33 -48 24—65 33—48 Общераспространенный прежде взгляд, что задержка солей в организме ребенка не зависит вовсе или зависит в весьма незначительной мере от поступления солей в организм, а регулируется только потребностями самого организма, в частности его ростом, в наст, время может считаться более или менее опровергнутым. Из приведенных выше цифр Ромингера видно, что у рожковых детей и детей, находящихся на смешан, вскармливании, абсолютное количество солей, задерживаемых в организме, значительно больше, чем у детей, получающих только грудь. У ребенка на искусственном вскармливании особенно много задерживается солей натрия и калия. Процент использования солей в нек-рых случаях повидимому у рожковых детей несколько меньше, чем у грудных. Повышенная задеряжа минеральных солей при искусственном вскармливании однако не ведет к гиперминерализации тканей, т. к. за периодами повышенной задержки солей следуют периоды усиленного их выделения и может быть даже непродолжительные периоды отрицательного баланса. Баланс отдельных солей дает значительные колебания в зависимости от возраста ребенка и его пищи и отчасти может быть от времени года. Часть всосавшихся солей (Na, К, соляной и фосфорной к-т) выделяется почками и выводится с мочой, другая часть (соли Са, Mg и Fe), пройдя через кровь, выделяется обратно в кишечник и выводится со стулом из организма. Последнее обстоятельство делает изучение минерального обмена особенно затруднительным. Потребность растущего организма в кальции меняется в зависимости от возраста ребенка, характера принимаемой им пищи и времени года. У здоровых, естественно вскармливаемых детей первого года жизни ежедневная потребность в СаО колеблется от 0,13 до 0,21 г, в среднем около 0,174г (Орглер); ребенку 6—7 лет для обеспечения нормального роста его костей необходимо ежедневно вводить 0,3—0,5 з СаО, из к-рых в организме задерживается около 30% (Herbst); ребенок 14 лет задерживает ежедневно около 0,45 з СаО, что возможно при суточном поступлении Са с пищей в количестве не менее 1,35 з. Э. и Ф. Мюллеры для детей старшего возраста дают следующие средние цифры необходимого ежедневного поступления кальция с пищей: в возрасте 6—8 лет—0,57—0,68 з, 10—12 лет—0,71—0,92 г, 13—15 лет—0,79—1,02 г СаО. Количество кальция, всасывающегося в кишечнике, не поддается учету; коли- чество кальция, задерживающегося в организме, меняется в зависимости от возраста и пищи ребенка; при грудном вскармливании, по Вангу (Wang), задерживается до 63%, при искусственном вскармливании до 47,5% и наконец у детей более старших—около 30% (Гербст). Около 5—10% введенного с пищей кальция выделяется с мочой, остальное выводится со стулом. В сыворотке крови кальция содержится около 10,5 мз%, из к-рых 2 мг в ионизированном виде, остальные приходятся на кальций органических соединений (около 25—30%) и на недиссо-циированные соли (о5—75%). Избыточное введение с пищей белков и жиров повидимому несколько ухудшает обмен кальция. Потребность растущего организма в фосфоре при искусственном вскармливании значительно больше, чем при естественном; в первом случае ежедневно должно вводиться с пищей 1—2 г Р205, во втором—0,2—Ю,3 г; у грудного ребенка задеряшвается 0,1—0,17 з (около 70%), у рожкового 0,23—0,5 (около 30%). У детей 6—8 лет ежедневная задержка Р205 на 1 кз веса равна 0,04 з, в возрасте 10—12 лет—0,034 з и в 13—15 лет—0,031 г (Мюллеры Э. и Ф.), что приблизительно составляет около 32—35% общего количества фосфатов, вводимых с пищей. Избыток введенных фосфорных соединений выделяется с мочой и стулом. В сыворотке крови соединения фосфора содержатся в виде неорганического (в среднем около 5,4 мг% Р205) и органич. Р; снижение фосфатов в крови ранней весной и нарастание поздним летом находятся в зависимости от богатства солнечного спектра ультрафиолетовыми лучами.— Магнезиальный обмен у детей изучен очень мало; у грудных детей удерживается 37,2% MgO, у рожковых— 11,5% (Langstein, Meyer), у детей более старших—от 14% до 49% (наблюдения Гербста на 4 детях). По наблюдениям Шлос-са (Schloss) над рахитиками как-будто существует антагонизм между известковым и магнезиальным обменом. Большая часть неусвоенного магния выделяется с мочой, меньшая—толстыми кишками. Магний понижает возбудимость нервной системы. Натрий и калий. У грудных детей (2—4 месяцев) за сутки задерживается Kj.0 — 0,17 — 0,19 з, у рожковых — 0,21— 0,28 з; Na20 в этом же возрасте при вскармливании грудным молоком удерживается 0,21 з, у ребенка, получающего коровье молоко,—0,22—0,44 з (Лангштейн-Мейер); у детей старшего возраста удерживается 2— 12,2% общего количества К20, введенного с пищей, a Na20—19,2—23%, что при пересчете на 1 кз "веса дает 0,002—0,009 К20 и 0,027—0,032 Na20 (Гербст). Неиспользованные щелочи у здоровых детей выделяются преимущественно с мочой, у больных значительно нарастает выделение их со стулом.—X лор. В цельной крови хлоридов содержится 450—550 мг%, в плазме— 570—620 мг%; но эти цифры подвержены значительным колебаниям в зависимости от состояния секреторной способности желудка. Точных данных, характеризующих обмен хлоридов у детей разных возрастов, да наст, времени нет. У детей старшего возра~ ста потребность не превышает 1—2 г. Выделение хлора происходит почти исключительно с мочой; суточное количество хлоридов мочи у грудных детей—0,1—0,14 г на 1 кг веса, на втором году—0,45, к 6 годам— 0,66, а после 10 лет—0,5—0,57 (Шанявский). Количество хлоридов в моче колеблется в зависимости от конституциональных особенностей ребенка (Мац).—Ж е л е з о. В женском молоке содержится около 1,4 мг°100, в коровьем—0,4—0,7 мг° 00 Fe203, при грудном вскармливании железа всасывается до 80,2%, при искусственном кормлении—28— 33% (Красногорский). Об обмене железа у детей старшего возраста данных не имеется. Железо необходимо для образования гемоглобина и кроме того, по Варбургу '.Warburg), играет роль катализатора при окислительных процессах. Органом выделения железа являются толстые кишки, в моче оно находится только в виде следов.—С ера поступает в организм глг-вным образом в виде цистина и в меньшем количестве в форме неорганических сернокислых солей. Потребность в сульфатах в грудном периоде— 0,022, на втором году жизни—0,056, на третьем году—0,075 и 0,06—0,07 на 1 кг веса в последующие годы. По Шварцу (Schwarz), ребенок пяти лет из введенных за сутки 0,103 г S03 выделил со стулом 0,008, с мочой—0,063 и удержал 0,032 г, что составляет 31 % поступления. Выделяется сера главным образом с мочой в виде неорганических сульфатов. Водный обмен. Значение водного обмена особенно велико для растущего организма ребенка. Ткани ребенка очень богаты водой: у 6-недельного плода 97,5% всей массы тела приходится на воду, у 5-месячного количество ее уменьшается до 90,^%, у семимесячного до 82,6% (Фелинг) и наконец ко времени рождения содержание воды в тканях падает до 68—70% (у взрослого около 60%). В течение всей дальнейшей жизни ребенка, начиная с 3—4-мес. возраста, происходит постепенное высыхание его тканей. Ткани ребенка не только богаты водой, но кроме того она в них находится в состоянии крайне неустойчивого равновесия (Ромингер, Лангштейн и Мейер). Эта гидролябильность, свойственная ребенку вообще, особенно резко выражена у детей конституционально аномалийных—эксуда-тиков, пастозных, лимфатиков. Количество воды, необходимое для правильного развития ребенка, подвержено значительным колебаниям в зависимости от возраста, совершаемой им работы, пищи, t° и влажности воздуха, конституции и пола. Потребность в воде в первые недели жизни, по Мейеру и Видмеру (Widmer), достигает 150—170 г, к 6 месяцам—110 гик концу года около 90 г на 1 кг веса. Приблизительно такие же цифры дает Пантелеева; по Шабановой, потребность в воде у старших детей постепенно снижается: с 95 г на 1 кг веса на втором году жизни и до 40 г к 13 годам. Ребенок годовалый за сутки должен получать около 800 г воды (учитывается вода, содержащаяся не только в жидкой, но и в твердой пище) (Czerny), ребенок 2—4 лет дохжен получать 957 г, 5—6 лет—1 200 г, 7—10 лет— 1 333 г, 11—14 лет—1 510 г. Потребность в воде у девочек несколько меньше, чем у мальчиков. Резорпция воды из кишечника у грудных детей в 3 раза больше, чем у взрослых, прохождение воды через жел.-киш. канал значительно ускорено, гидремическая реакция при водной пробе у детей наступает быстрее, водное равновесие после сухоядения у взрослых выравнивается только через 5 часов, а у детей через 1V2 часа (Ромингер). Все это говорит о крайне напряженном водном обмене у детей; Мейер и Нассау (Meyer, Nassau) указывают, что каждая частица воды за время' прохождения через организм ребенка не менее 5 раз совершает путь: кишечник—сосуды—кишечник. По Каммереру, у грудного ребенка 59—60% воды удаляется почками, 33% кожей и легкими, 6% со стулом и только 1—2% задерживается в организме. Распределение выделяемой воды может сильно меняться у одного и того же ребенка в зависимости от внешних условий и принимаемой ими пищи. Экстраренальное выделение воды (через кожу и легкие), или т. н. perspiratio insensibilis, у детей относительно больше, чем у взрослых, и лишь несколько понижено в период новорожденно-сти. По Бирку и Эделыптейну (Birk, Edel-stein), новорожденный выделяет этим путем в течение суток 21,7—37,5 г на 1 кг веса; нормальный грудной ребенок выводит экс-траренально 32,2—38,3 г на 1 кг веса (pro die), рожковый ребенок—44,4 г (Heubner, Rubner). По Каммереру, путем perspiratio insensibilis за сутки выделяется у ребенка первых дней около 80 г, к концу 1-й недели—100—110 г, на 4-й неделе—130 г, на 10-й—180 г, к полугодию—около 300—400 г и к концу года—около 500 г. После года экстраренальное выделение нарастает медленно, составляя на 10-м году 550—650 г, а к 11 -IP/a годам 680—780 г. Perspiratio insensibilis в бодрствовании больше, чем во сне, при беспокойстве и крике больше, чем в покое, в вертикальном положении больше, чем в горизонтальном и наконец у детей-дистрофиков (27—40%) больше, чем у эйтрофиков (33—60%) (Локшина и Александрова). Водный обмен стоит в тесной связи с углеводным и минеральным. Выключение углеводов из пищи ребенка вызывает падение веса, и, наоборот, введение их способствует задержке воды. Кальций и калий усиливают отдачу воды тканями, NaCl—в одних случаях содействует задержке воды в организме, в других случаях может усиливать ее выделение. При концентрированной пище диурез уменьшается, а экстраренальное отделение возрастает (Бош). Водный обмен в значительной мере находится в зависимости от состояния эндокринной системы ребенка. Ограничение количества вводимой воды ведет к явлениям эксикоза: западение родничка, падение тургора тканей, сухость слизистых, мозговые явления, концентрированная моча и сухой темный стул. Ограничение воды при достаточном введении белков вызывает явления еще более тяжелые, напоминающие картину пищевой интоксикации, и сопровождается повышением 1°тела 76ft (Мюллер, Ричель, Финкелынтейн). Изменения со стороны обмена веществ при патологических состояниях (болезни обмена) и аномалиях конституции—см. соответствующие слова. а. Тур. Пигментный обмен, см. Пигменты. Лит.: Бакшт Г., Беременность и обмен веществ, М.—Л., 1929; Богомолец А., О вегетативных центрах обмена, М., 1928; Гефтер А., Основной обмен и его значение в клинике внутренних заболеваний, Клин, мед., 1930, № 2; Д го б у а Е., Обмен веществ с точки зрения практического врача, М.—Л., 1927; 3 ю к о в А., Обмен воды в организме, Харьков, 1929; Лаббе М. и Непвб Ф., Ацидоз и алкалоз, М.—Л., 1931; Николаев Н., Обмен веществ и проблема кроветворения, М., 1927; о н ж е, Руководство по межуючному обмену, М., 1930; Н о-орден К. и Шиттенгельм А., Терапия болезней обмена, М.—Л., 1930; Р у б и н ш т ей н Д., Основной обмен и клиническое значение определения его, Вестн. совр. мед., 1927, № 10; Скворцов В., Ацидоз и алкг лоз в медицине, М., 1928; У м б е р Ф., Болезни обмена веществ,М.—Л., 1929; Фромгольд Б., Болезни обмена веществ (Частная патология и терапия, под ред. Г. Ланга и Д. Плетнева, т. IV, в. 1, М.—Л., 1928); Цунтц Л., Обмен веществ женщины, М., 1928; А с h а г d Ch., Troubles des echanges nut-ritifs, v. I—II, P., 1926; Boenlieim F., Wasser- u. Minerals^ ITwechsel u.innereSekretion, Halle a./S-,1927; Jturnal of metabolic research, v. I—"VIII, Morristown, 1922—26; Grafe E., Die pathologische Physiologie des "Gesamtstoff- u. Kraftwechsels bei der Erna.hr ung des Menschen, Erg. d. Physiol., B. XXI, T. 2, 1923 (также отд. изд.—Miinchen, 1923); он же, Die Krank-heiten des St< ffwechsels u. ihre Behnndlung, B.—Wien, 1931; Handbuch der Biochemie des Menschen u. der Tiere, hrsg. v. С Oppenheimer, B. V—IX, Jena, 1925— 27 (лит.); Handbuch der biologischen Arbeitsmethoden, hrsg. v. E. Abderhalden, Abt. 4, T. 9—Methoden der quantitativen Bestimmungdes Stoffwechsels des Gesamt-organismus, von Organen u. Zellen, В.-—Wien, с 1923; Handbuch der ш rmalen und pathob gischen Physiologie, hrsg. v. G. Bergmann, A. Bethe u. a., B. II, III, V, XVI, XVII, В., 1925—31 (монографическое изложение, лит.): Handbuch derPathologie des Stoffwechsels, hrsg. v. С Noorden, В., 1906; К nipping H. u. Kowitz H., Klinische Gasstoffwechseltechnik, В., 1928; L a b b ё M. et S L.e v e n i n H., Le metabo-lisme basal, P., 1929; L a b b ё M. et V i о 1 1 e P., Metabolisme de l'e^u, P., 1927; L i с h t w i t z L., Stoffwechselkrankheiten (Hndb. d. inneren Medizin, hrsg. v. G. Bergmann u. R. Staehelin, B. IV, T. 2, B.,. 1926, лит.); Lusk G., The elements of the science of nutrition, Philadelphia, 1928; L о е w у A., Gas-wechsel u. Eiweisstoffwechsel unter dem Einfluss der inneren Sekretion, Ls z, 1927; Macleod J., Kohlen-hydratsttffwechsel u. Insulin, В., 1927; Spezielle Pa-thtlogie u. Therapie innerer Krankheiten, hrsg. V. F. Kraus u. Th. Brugsch, B. I—Erkrankungen des Sti ffwechsels, B.—Wien, 1919 (лит.); Strauss H., Kohlenhydrat-Stoffwechsel u. innere Sekretion, Halle, 1927; Tern ine E. et Z u n z E., Le metabolisme de base, P., 1925 (лит.); ThannhauserS., Stol'f-wechsel u. Stoffwechselkrankheiten, Munchen, 1929. Обмен веществ у детей.—Г ельмрейх Э., Обмен энергии у ребенка, М.—Л., 1928; Л у н ц Р., Физиология и диететика грудного ребенка, М.—Л., 3-е изд., 1930; Маслов М.,, Основы учения о ребенке, 3-е изд., т. I, М.—Л., 1930; В i r k W., Untcrsuchun-gen uber den Stoffwechsel des Kindes im Fieber, В., 1926; Helmreich E., Besonderheiten des Kraftwechsels im Kindesalter, Klin. Wochen-schr., 1929, № 35; о н ж e, Der Grundumsatz im Kindesalter, Erg. d. inn. Med., B. XXXV, 1929; Knauer H., Ergeb-nisse der Lipoidst ffwechselforschung mit bes. Beruck-sicbtigung des Kindesalters, В., 1928 (лит.);М ii 1 1 e г E., Stoffwechsel u. Ernahrung alterer Kinder (Hndb. d. Kinderheilkunde.'hrsg. v. M. Pfaundler u. A. Schloss-mann, B. I, Lpz., 1923; поел. изд.—1931); Schiff E., Das Exsiccoserroblem, В., 1929. См. также лит. к ст. Грудной ребенок. (metabolisme intermediaire, intermediarer Stoffwechsel), термин, предложенный для обозначения превращений, испытываемых введенными в организм питательными веществами от момента их всасывания в кишечнике до использования в организме,' resp. их выделения почками, кожей и легкими (см. Обмен веществ). Смотрите также:
Смотрите также:
- ОБМОРОК, обмирание, synkope, состояние б. ч. непродолжительной полной потери сознания с падением деятельности сердца и ослаблением дыхательных движений. О. наступает б. ч. внезапно, но иногда следует за состоянием дурноты, т. ...
- ОБОИ служат для отделки и украшения внутренних стен жилищ. О. в Европу проникли из Китая; первые обойные фабрики появились в Англии в 18 в. Выработка О. в основном представляет собой выработку ...
- ОБОЛОЧКА (оболочка клеток). Протоплазма всякой клетки, растительной или животной, отграничена от окружающей среды более плотным слоем. Последний может быть очень тонок, почти незаметен, но может развиваться и ...
- ОБОНЯНИЕ, физ.-хим. процесс, при помощи к-рого человек и животное получают представление о запахе. Особо важную роль О. играет у животных и у нек-рых из них О. (син.' чутье) очень велико; ...
- ОБОНЯТЕЛЬНЫЙ НЕРВ, см. Olfactorius nervus.