АРХИТЕКТОНИКА КОРЫ ГОЛОВНОГО
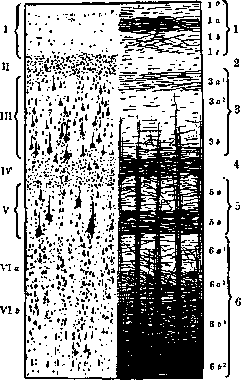
Рисунок 1. Слева — шесть основных клеточных слоев коры по Brodmann'y: I—lamina zonalis, молекулярный или тангенциальный слой; //— 1. granularis externa, наружный зернистый слой; III—1. pyramidalis, пирамидальный слой; IV—1. granularis interna, внутренний зернистый слой; V—1. ganglionaris, ганглиоз-ный слой, или слой глубоких пирамид; VI а и b—lamina, multiformis, полиморфный слой, распадающийся на слой треугольных клеток (Via) и слой веретенообразных клеток (VIb). Справ а—миэлоархитектоническая картина по Vogt'y: 1—1. tangentialis с подразделением на sublamina superficialis (i°), subl. intermedia с двумя частями—pars externa (la) и pars interna (lb) и sub), profunda (lc); 2—1. dysfi-brosa; 3—1. suprastriata с тремя sublamina: subl. superficialis (За1), или stria Kaes-Bech-terewi, subl. intermedia (За2) и subl. protunda (3b); 4—stria Baillargeri externa, или наружная поперечная полоска; 5—1. interstriata (5а) » и stria Baillargeri interna (Sb), или внутренняя
поперечная полоска; 6—1. substriata («а1, ва'), 1. limitans interna (вЬг) и album gyrorum (eb'). В нек-рых частях коры шестислойный клеточный тип не выражен, в других он как бы недоразвит и в третьих он подвергается различным модификациям. Каждый слой в отдельности может оказаться то шире, то уже; нервные клетки его то большей, то меньшей величины; слой их то гуще, то реже; кроме того, и располагаются они по-разному. Иногда основной слой бы распадается на sublaminae. Основные слои могут и совсем исчезать. Так, в area gigantopyramidalis исчезает lam. granularis interna; в area striata она расслаивается на три слоя. Рисунок 1 показывает, что миэлоархитектоника дает еще более детальное деление коры на слои, чем цитоархитектоника. Разница в строении ми-элоархитектонических картин может выразиться в следующем: лучевые (радиальные) волокна могут быть разной толщины, а пучки их—разной густоты; длина их тоже может варьировать; поперечные волокна отличаются то толщиной, то образованием различных пластов. Эмбриологические основы. Биол. значение шестислойного типа коры и ее вариаций- выяснено эмбриол. исследованиями Бродмана и дополнено последними исследованиями Розе. К концу третьей недели развития человеческого зародыша передний отдел мозговой трубки распадается на пять пузырей, из к-рых самый крайний—конечный мозг—дает полушария. Начальная эпителиальная структура стенки полушария сначала преобразуется в миэлоспонгий, состоящий из спонгиобластов и невробластов (см. рисунок 2),—зародышевый пласт. Затем идет расслоение его на matrix и краевой слой (средний срез рис. 2), а из matrix диференци-руется пирамидный или корковый слой (Ру, нижний срез рис. 2). Пирамидный слой растет и утолщается, и на 5-м месяце утробной жизни его невробласты начинают группироваться послойно. Первоначально, на 6-м мес. утробной жизни, обособляются 5-й и 6-й слои. Затем, на протяжении от 6-го до 8-го месяца, из общей клеточной массы выделяются и остальные слои. Т. о., разновременно—в одном месте коры раньше, в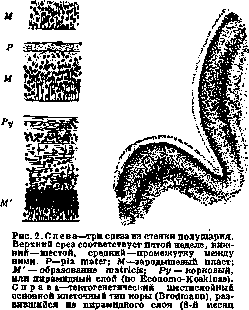
утробной жизни).
другом несколько позже — появляется тек-тогенетическая шестислойная кора. Но она формируется не всюду: есть участки коры большого мозга, к-рые через этот стадий развития не проходят, т. е. в них шесть слоев или не образуются или они недоразвиваются Наружная поверхность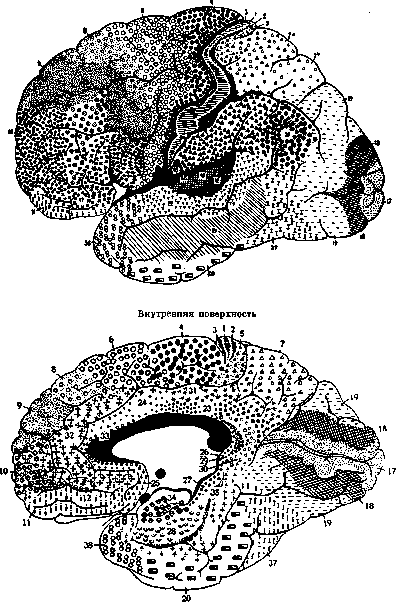
Рисунок 3. Ареальная карта Brodmann'a. Названия полей: 1—a. postcentralis Intermedia; 2—a. postcentralis caudalis; 3—a. postcentralis oralis; 4—a. prae-centralis gigantopyramydalis; 5—a. praeparietalis; 6—a. frontalis agranularis; 7—a. parietalis superior; 8—a. trontalis intermedia; 9—a. frontalis granuiaris; 10—a. frontopolaris; 11—a. praefrontalis; 17—a. striata; J*—a. occipitalis; 19—a. praeoccipitalis; 20—a. temp oralis infer.; 21—a. tempor. media; 22—a. temper. sup.; 23—a. limbica post, ventralis; 24—a. limb. ant. ventr.; 26—a. subge-nualis; 26—a. ectosplenialis; 27—a. praesubicularis; 28—a. entorhinalis ventralis; 29—a. retrolimbica granuiaris; 30—a. retrolimbica agranularis; 31—a. limbica post, dorsalis; 32—a. limb. ant. dors.; 33—a. praegranularis; 34—a. entorhinalis dors.; 35—a. perirhinalis; 36—a. ectorhinalis; 37—a. occipito-temporalis; 38—a. temporopolaris; 39—a. pariet. inf. post. s. angularis; 40—a. pariet. inf. ant. s. supragranularis; 41—a. tempor. transversa ant.; 42—a. temp, transv. post.; 43 —a. subcentralis; 44 — a. opercularis; 45 — a. triangularis; 46 — a. frontalis media; 4 7—a. orbitalis.
(гетерогенетические формации). Такова филогенетически самая старая кора большого мозга (см. ниже). В новой коре шестислой-ный этап развития является обязательным для всех ее частей (гомогенетические формации); однако, по своему значению шесть слоев здесь не тождественны шестислойной коре зрелого мозга. Это лишь первичная шестислойная клеточная закладка, подлежащая дальнейшей диференцировке. Дифе-ренцировка же ее идет не сразу, а отдельными участками коры. На нек-рых из них шестислойный тип сохраняется и в дальнейшем (гомотипические формации), на других число основных слоев или сокращается или умножается (гетеротипические формации). Появляются и другие особенности строения. Так, постепенно формируются отдельные поля—агеае—с теми структурными признаками, которые определяются в зрелом мозгу при архитектонических исследованиях. Сравнительно-анат. параллели. Сравнительная анатомия учит, что на разных уровнях филогенетического развития производные конечного мозга, в частности полушария, далеко не равноценны. Их структура и функции у рыб и амфибий тесно связаны с обонятельной областью. Но уже у амфибий в этой старейшей коре (archipalli-um) обособляются два отдела: palaeocortex, связанный с bulbus olfactorius, и archicor-tex—будущий Аммонов рог. Далее, на еще более высокой ступени развития—у рептилий—начинает развиваться новая кора (neopallium, neocortex), достигающая у высших позвоночных громадного развития. Как сказано выше, у человека archipallium, т.е. старая обонятельная кора, и в процессе онтогенетического развития имеет свой особый тип развития: онане проходит стадия шестислойной коры. Для neopallium (neocortex) этот стадий обязателен и характерен. Старая и новая кора в зрелом мозгу также имеют и значительные структурные особенности. О. Фохтом установлено, что лучевые волокна в новой коре достигают или до верхней границы 5Ъ (тип infraradiatus) или до границы между ЗЪ и Зс (тип intradiatus). По его терминологии, это isocortex—часть коры, соответствующая neocortex. В старой коре лучевые волокна доходят до верхнего отдела третьего слоя (тип supraradiatus). Он называет ее allocortex. В последнее время О. Фохт, на основании исследования Розе и Яна (Jan), отнес infraradiatus к allocortex. Ламинарная структура и функция. При изучении послойной, или ламинарной структуры коры приходится различать, собственно, строение слоев, их число, состав каждого слоя и пр. Соответствующие данные, касающиеся отдельных area, настолько разнообразны и обширны, что могут быть изложены только при наличии иллюстрационного материала. Необходимо лишь отметить, что миэлоархитектониче-ский метод, как видно из рис. 1, позволяет делить кору в ламинарном отношении еще более детально, чем метод цитоархитекто-нический. Благодаря этому, получается и бблыпая ареальная диференцировка коры по этому методу, отсюда же и большее число полей (Фохт). Кроме состава слоев, изу- чается (лучше всего по методу Golgi) и связь между ними (синаптология). Многое сделано в этом отношении Рамон-и-Ка-халом (S. Ramon у Cajal). Но связь между слоями и соседними полями исследуется также экспериментально, путем проводимых в разных направлениях разрезов, вставления изоляционных пластинок, раздражения током и пр. Что касается ламинарной функции, то в отношении ее нет полного единодушия. По Капперсу (Kappers), в старой коре (palaeocortex) имеются два взаимно связанных главных слоя: поверхностный— слой зерен и глубокий—пирамидный слой. Клетки первого имеют рецепторную функцию, второго — проекционно-комиссураль-ную и ассоциативную. В новой коре (neocortex), в свою очередь, можно различать два отдела или зоны,только более развитые: наружную, в к-рую входят наружные слои, начиная с четвертого, и внутреннюю, состоящую из пятого и шестого слоев. Тот же автор устанавливает, что наружная зона генетически и морфологически соответствует зернистому, а глубокая зона—пирамидному слою старой коры. И в фнкц. отношении между ними имеются аналогии. Рядом опытов доказано, что глубокая зона neocorticis относительно независима от поверхностной. Кортико-спинальные, кортико-бульбарные, кортико-понтинные, кортико-таламические волокна имеют источником клетки глубокой инфрагранулярной зоны. Оттуда же исходят волокна мозолистого тела. Наоборот, в супрагранулярную зону идут афферентные волокна. Особенно много их в сенсорных центрах, где сильно развит также слой внутренних зерен. В наружной зоне начинаются также ассоциативные волокна, связующие между собой и интрагризеально и экстрагризеально отдельные части коры. Т. о., внутренняя зона является, по преимуществу, отправной станцией для переноса невродинамической энергии; наружные же слои клеток, гл. образ., выполняют рецепторную и ассоциативную функции. Ареальная структура коры. Генетический принцип диференцировки коры не целиком, а отдельными участками, касается и старой и новой коры. В результате, всюду в зрелой коре мы находим развившиеся по общим законам органообразова-ния гетеропластические, т. е. составленные из разных структурных элементов, поля— агеае. Весьма характерно в построении их то, что морфологическая граница каждого поля имеет линейную форму. Она только кое-где совпадает с бороздами мозга, но может итти и по выпуклой поверхности извилин. Агеае имеются и в коре различных млекопитающих. Сравнивая между собой ареальную структуру коры большого мозга у различных млекопитающих, можно установить следующее: 1) общее расположение полей имеет сегментарный характер, 2) есть поля, к-рые сохраняют одно и то же послойное строение и занимают в коре одни и те же места у всех этих животных, 3) у всех млекопитающих выявляются сходные по структуре и расположению комплексы полей, так наз., области. У низших из этих позвоночных полей меньше и они менее Наружная иоверхность ~-4&Z /^ и§ Г 4i Plm^s / у— [ '***Ш5ш1 * jf ^Щ Pin D«mi ^V /Гит/ V те^ШШ / ^S2 ^EL psp X /у «А к™ JSmE, If /"&&--■-■■ ^-/ШШ$ IwPi JfellsSp^V^Pg^ ШШтМВ^^Е^^^т^Ж^Ж^Л LzMI9mm 11Ш Hr^ jffl^^PS^^^^^^^i «fjtwwlm гг^^^^^^И г*„* JIlllMt Ир! Внутренняя поверхность tia | га г£—-*|*ЯК>>^ JVft) Ре f Л#8ЯЙ£> : О^Ц§£^ Bltf \Рв« /^ч**вйИ ^e^siS?^ ш?Ш F^ \& / i*^?№iap5fe3gij3sSg НИШ» к2#^^^^^^ v\ l?j та illl^si >HJ„ нвЛ ^^^^"7 X^^^^^fc Шы?»ж --Жч'У* ^~^Г~ "^T^LL^r}'^^ ££0 ЙЖ^^У^ ЕЛйЙКд Ш111 ШшШк W$F* Низ Hi, н«* Рисунок 4. Ареальные карты Economo и Koskinas (обозначения—см. след. стр., рис. 5). диференцированы. Наивысшей сложности ареальная структура достигает у человека. При этом разница в тонкой ареальной структуре между мозгом человека и высшими антропоидными обезьянами оказывается гораздо большей, чем между антропоидными и низшими обезьянами [вопреки закону Гёксли (Huxley)]. Впрочем, есть области в коре человека, которые менее развиты у человека, чем у других животных. Деление коры на области. На основании нек-рых общих морфологических признаков часть полей можно выделить в один общий комплекс—область, при чем вся кора распадается на несколько таких областей (regiones). Области встречаются у всех млекопитающих, что указывает на нек-рую постоянную и общую для каждой из них функцию. Бродман различает Ш областей: I) г. postcentralis (включает поля задней центральной извилины 1, 2, 3, 43); 2) г. praecentralis (включает поля передней центральной извилины 4, 6); 3) г. fronta-Jis—все поля лобной доли кпереди от г. praecentralis и cingularis; 4) г. insularis— область островка; 5) г. parietalis—поля 5, 7, 39, 40; 6) г. temporalis—поля височной доли; 7) г. occipitalis—затылочная область— поля 17, 18, 19; 8) г. cingularis—поля 23, 24, 25, 31, 32, 33; 9) г. retrosplenialis—поля 26, 29, 30; 10) г. iippocampica—поля 27, 28, 34, 35; 11) г. olfactoria—обонятельная область.—Экономо и Коскинас предлагают несколько другое деление на области и lobi. Функциональное значение областей и полей. Несмотря на кажущуюся Нижняя поверхность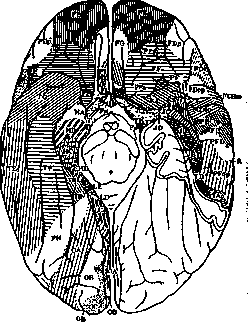
Верхняя поверхность FE
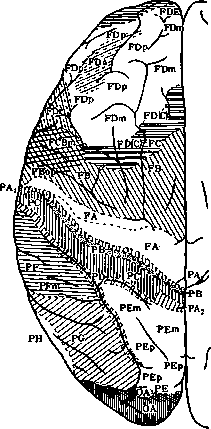
Рисунок 5. Ареальиые карты Economo и Koskinas. Regio praerolandica: area praecentralis (FA); a. Irontalis agranularis (FB); a. frontalis intermedia (FC) и др. Regio frontalis: a. frontalis granularis (FB); a. frontopolaris (FE) и др. Regio orbitalis: a. orbitalis (granularis) (FF); a. gyri recti (FG); a. praefrontalis (FH); a. frontoinsularis (FT); a. pi-riformis frontalis (FK); a. parolfactoria (FL); a. geniculata (FM); a. praecommissuralis (FN). Hegio limbica sup. ant.; a. limb. ant. agr. {LA); a. ultracingularis ant. (LB,); a. indusei (.LB,). Regio limb. sup. post.: a. cingularis post, dorsalis (LC,); a. cingularis post, ventral. (LCS); a. cingularis lim. post. (LC,). Subregio retrosplenialis; a. retrosplenial. agranularis (LD); a. retrosplemalis granulosa (LE); a. ultracingularis post. (LFJ; a. (ultracingularis) obtecta (LF2). Lobus insulae: a. insul. praecentralis (JA); a. insulae postcentralis (JB); a. orbito-insularis (JC); a. insularis piriformis (JD). Regio postcentralis (parietalis ant.): a. postcentralis gigantopyrami-dalis (PAJ; a. postcentralis oralis simplex (PB); a. postcentr. intermedia (PC); a. postcentr. caudalis (PD). Regio parietalis sup.: a. parietal, sup. (PE). Regio parietal, inf.: a. supramarginalis (PF); a. angularis (PG). Regio parietalis basalis: a. pariet. (temporo-occipitalis) basalis (PH). Lobus occipitalis: a. peristriata (OA); a. parastriata (OB); a. striata (granulosa) (ОС). Regio supratemporalis: a. temp. sup. (ТА); a. supratempor. magnoc. simplex (ТВ); a. supratemporalis granulosa (TC); a. supratemp. intercalata (TD). Regio tempor. propria: a. tempor. propria (ТЕ). Regio fusiformis: a. fusiformis (TF); a. hippocam-potemporalis (TH). Regio temporopolaris: a. temporopolaris (TG); a. piriformis tempor. (TG); a. subst. pert. post. (TH). Lobus limbicus inferior: a. uncinata (HA); a. parauncinata (ЯВ); a. rhinalis limitans (HC); a. praesubicular. granul. (HD); a. pyramidalis (HE); a. fasciae dentatae (HF).
обособленность клеток полей при распределении их по слоям, в действительности они тесно сплетаются друг с другом своими отростками и образуют один неврональный комплекс. В этом комплексе каждая своеобразно диференцированиая клетка или каждый слой имеют свои биол. функции, и все они, в свою очередь, покрываются нек-рой биологической же функцией ареального целого. Данные физиологии о локализации в коре отдельных отправлений мозга в общем соответствуют делению коры на области и поля. Чтобы убедиться в этом, достаточно сравнить ареальные и локализационные карты. При этом ни у кого не возникает сомнения в том, что нек-рые проявления мозговой деятельности локализуются в тех или других областях. Спорным является только вопрос о том, совпадает ли какая-либо мозговая функция точно с пределами той или другой агеае. Проблема эта сложна и трудна для решения. Многие проявления мозговой деятельности в действительности не так просты, как кажутся, и соответствуют часто деятельности целого мозга или его части, но все же есть другие функции^—более простые и более локализованные. Нек-рые из них, несомненно, соответствуют area. Имеют большое значение в этом отношении данные работ Геншена, Вильбранда, Минковского и Монакова (Henschen, Willbrand, Minkow-sky, Monakow), которые позволяют связывать элементарный акт зрения с областью fiss. calcarinae (area striata). Показательны и новые опыты С. и О. Фохтов с электро-раздрая:имой корой церкопитека, при чем оказалось,что каждому полю,предварительно обособленному на основании цитоархи-тектоники, соответствует особая функция. Особенно поразительно совпадение по сходству двух ареальных карт—одной, составленной О. Фохтом для человеческого мозга по данным эксперимента над церкопитеком, и другой, составленной Ферстером (Foerster) для человеческого же мозга на основании раздражения коры при операциях. ; А. и высшая нервная (психическая) деятельность. Архитектоника коры головного(болыного)мозга как органа •высшей нервной (психической) деятельности имеет большое значение и для исследования этой последней. Так, архитектоника дает руководящие указания и рефлексологу, занимающемуся образованием условных рефлексов, и психологу, изучающему поведение животных: и тот и другой, производя добавочно экспериментальное раздражение Или разрушение коры животных, имеют воз-|можность базироваться при этом не только да топографии борозд и долей, но и на тонкой ареальной структуре. Новое слово вносит далее архитектоника коры головного мозга в изучение того естественного эксперимента над человеком, каким является душевная болезнь. Патологическая анатомия т. н. дефективных психозов (слабоумий) в свете топистики, патоклизы и патоархитектоники (С. и О. Фохты) получила уже новое понимание. При этом наглядно выявился ;анат. субстрат и идиотии и приобретенных интеллектуальных дефектов. Ценность архитектонических исследований надо считать доказанной и по отношению к здоровой психике. Имеется ряд микроскопических исследований, устанавливающих особенности индивидуальной архитектоники. Намечаются архитектоника расовая, архитектоника преступников и т. д. Особенно большое значение приобретает изучение архитектонических особенностей мозга высокоодаренных людей. В этих исследованиях важны не только получаемые новые морфологические данные, но еще в большей степени установление соотношений между ними и теми или Другими психическими особенностями данных лиц. Задача будущего не только точно установить такие соотношения, но и доказать их закономерность, т. е. то, что они 'удовлетворяют математической формуле корреляций. Для этого, очевидно, понадобится произвести большое число исследований. В общем, можно сказать, что все данные произведенных до сих пор архитектонических исследований как на людях, так и на животных, подтверждают следующее общебиологическое положение: в процессе органического развития диференцировке формы соответствует диференцировка функции; психическая деятельность, представляющая собой одну из функций мозга, не составляет исключения из этого положения. Ареальные атлас и карты. Выше говорилось о трудности дать описание ламинарной структуры агеае без рисунка. Эта нужда восполняется фотографиями цито- и миэлоархитектонических картин каждой агеае. С ними каждый, занимающийся исследованием архитектоники коры голов- ного мозга, может сверять свои препараты при исследовании индивидуальных, расовых, возрастных и патологических особенностей строения коры. Из таких фотографий составляется атлас (см. атлас и точное описание каждой из area у Экономо-Коскинас). Раз определены границы отдельных агеае, то из них легко составить карту мозговой поверхности (см. рисунок 3, 4, 5). Патоархитектоника и па ток л и-з а. Применение архитектонических методов к изучению нервных и душевных б-ней привело С. и О. Фохтов к учению о патоар-хитектонике и патоклизе. По С. и О. Фох-там, диагносцировать то или другое страдание мозга можно часто на основании одной лишь патоархитектоники. Ими доказано это по отношению к болезням corporis striati, и они пытаются также распространять этот принцип на б-ни головного мозга. В литературе имеется ряд патоархитектони-ческих исследований различных дементных болезненных форм (дефект-психозов), латерального склероза, Гетчинсоновской хореи, церебрального паралича при целости пирамидных путей, раннего слабоумия, эпилепсии, прогрессивного паралича, маниакально-депрессивного психоза и др. Интересно, что изменения патоархитектоники идут по определенным законам.Многие вредности имеют сродство с отдельными невро-нальными комплексами (патоклиза), поражая их больше, чем другие. Напр., пора^ жается III и V слои или только IV слой. Это—ламинарная патоклиза. В меньшей степени проявляется она на отдельных агеае (ареальная патоклиза). Лит.: Betz w., tlber die feinere Struktur der Grosshirnrinde, Zentralblatt 1. med. Wiss., 1874 u. 1881; V о g t C. u. O., Allgemeine Ergebnisse unserer Hirnforschung, Jahrb. f. Psychologie u. Neurologie, 1919, B. XXV, Erganzungsheft; и х ж e, Erkran-kungen der Grosshirnrinde im Lichte der TopistiX, Pathoklise u. Pathoarchitektonik, 1922, Erganzungsheft; Brodmann K., Vergleichende Lokalisations-lehre der Grosshirnrinde, Leipzig, 1925; E со no mo С. и. К oskin as &., Die Zytoarchitektonik der Hirn-rinde des erwachsenen Menschen, Textband und Atlas, Wi en, 1925; Kappers A., Die vergleichende Anatomie des Nervensystems der Wirbeltiere und des Menschen, 1921; EoseM., ttber das histogenetische Prinzip der Einteilung d. Grosshirnrinde, Jahrbuch /Or Psychologie und Neurologie, 1926, Band XXXII; E с o-nomo C., Zellaufbau der Grosshirnrinde des Menschen, Berlin, 1927. П. Снесарев. Смотрите также:
Смотрите также:
- АРХОПЛАЗМА (от греч. arche—начало и plasma-—образование), син. архиплазма, термин, предложенный Бовери (Boveri) для обозначения вещества, из к-рого состоят нити веретена и т. н. лучи клеточного сияния, возникающие при митотическом делении ядра. ...
- АРХЭНТЕРОН (от греч. arche—начало и enteron—кишки), первичная кишка, полость гаструлы, выстланная энтодермой; наружу первичная кишка открывается первичным ртом (бластопором).
- АРХЭНЦЕФАЛОН (от греч. arche—начало и enkephalos—мозг), передняя часть эмбрионального первичного мозга позвоночных животных, из которой в дальнейшем развиваются передний мозг (prosencephalon) и средний мозг (mesoncephalon).
- АРЦБЕРГЕРА АППАРАТ (Arzberger), был предложен для лечения геморроя, а в даль-i^^^ нейшем и для лечения проста-■чЙп^^вХ ты с целью воздействия холо-Щ ...
- АСАПРОЛ, см. Абрастол.